
Actin - Actin
| Actin | |||||||||
|---|---|---|---|---|---|---|---|---|---|

Diagrama de fita de G-actina. O ADP ligado ao sítio ativo da actina (varas multicoloridas perto do centro da figura), bem como um dication de cálcio complexado (esfera verde) são destacados.
| |||||||||
| Identificadores | |||||||||
| Símbolo | Actin | ||||||||
| Pfam | PF00022 | ||||||||
| InterPro | IPR004000 | ||||||||
| PRÓSITO | PDOC00340 | ||||||||
| SCOP2 | 2btf / SCOPe / SUPFAM | ||||||||
| |||||||||
A actina é uma família de proteínas multifuncionais globulares que formam microfilamentos no citoesqueleto e os filamentos finos nas fibrilas musculares . É encontrado essencialmente em todas as células eucarióticas , onde pode estar presente em uma concentração de mais de 100 μM ; sua massa é de aproximadamente 42 kDa , com diâmetro de 4 a 7 nm.
Uma proteína actina é a subunidade monomérica de dois tipos de filamentos nas células: microfilamentos , um dos três principais componentes do citoesqueleto, e filamentos finos, parte do aparelho contrátil nas células musculares . Pode estar presente como um monômero livre chamado G-actina (globular) ou como parte de um microfilamento de polímero linear chamado F-actina (filamentoso), ambos essenciais para funções celulares importantes como a mobilidade e contração das células durante divisão celular .
A actina participa de muitos processos celulares importantes, incluindo contração muscular , motilidade celular , divisão celular e citocinese , movimento de vesículas e organelas , sinalização celular e estabelecimento e manutenção de junções celulares e forma celular. Muitos desses processos são mediados por interações extensas e íntimas da actina com as membranas celulares . Em vertebrados, foram identificados três grupos principais de isoformas de actina , alfa , beta e gama . As alfa actinas, encontradas nos tecidos musculares, são os principais constituintes do aparelho contrátil. As actinas beta e gama coexistem na maioria dos tipos de células como componentes do citoesqueleto e como mediadores da motilidade celular interna . Acredita-se que a diversidade de estruturas formadas pela actina, permitindo-lhe cumprir uma ampla gama de funções, seja regulada pela ligação da tropomiosina ao longo dos filamentos.
A capacidade de uma célula de formar microfilamentos dinamicamente fornece a estrutura que permite que ela se remodelar rapidamente em resposta ao seu ambiente ou aos sinais internos do organismo , por exemplo, para aumentar a absorção da membrana celular ou aumentar a adesão celular para formar o tecido celular . Outras enzimas ou organelas , como os cílios, podem ser ancoradas a esse arcabouço para controlar a deformação da membrana celular externa , o que permite a endocitose e a citocinese . Ele também pode produzir movimento por si só ou com a ajuda de motores moleculares . A actina, portanto, contribui para processos como o transporte intracelular de vesículas e organelas, bem como contração muscular e migração celular . Portanto, desempenha um papel importante na embriogênese , na cicatrização de feridas e na invasão das células cancerosas. A origem evolutiva da actina pode ser rastreada até células procarióticas , que possuem proteínas equivalentes. Homólogos de actina de procariotos e arquea polimerizam em diferentes filamentos helicoidais ou lineares que consistem em uma ou múltiplas fitas. No entanto, os contatos na fita e os locais de ligação de nucleotídeos são preservados em procariotos e em arquéias. Por último, a actina desempenha um papel importante no controle da expressão gênica .
Um grande número de doenças e enfermidades são causadas por mutações em alelos dos genes que regulam a produção de actina ou de suas proteínas associadas. A produção de actina também é fundamental para o processo de infecção por alguns microrganismos patogênicos . Mutações nos diferentes genes que regulam a produção de actina em humanos podem causar doenças musculares , variações no tamanho e na função do coração e surdez . A constituição do citoesqueleto também está relacionada à patogenicidade de bactérias e vírus intracelulares , principalmente nos processos relacionados à evasão das ações do sistema imunológico .
Descoberta e investigação inicial

A actina foi observada pela primeira vez experimentalmente em 1887 por WD Halliburton , que extraiu uma proteína do músculo que 'coagulou' preparações de miosina que ele chamou de "fermento de miosina". No entanto, Halliburton foi incapaz de refinar ainda mais suas descobertas, e a descoberta da actina é creditada a Brunó Ferenc Straub , um jovem bioquímico que trabalhava no laboratório de Albert Szent-Györgyi no Instituto de Química Médica da Universidade de Szeged , Hungria .
Seguindo a descoberta de Ilona Banga & Szent-Györgyi em 1941 que a coagulação só ocorre em algumas extrações de misosina e foi revertida com a adição de ATP, Straub identificou e purificou a actina das preparações de miosina que coagularam. Com base no método de extração original de Banga, ele desenvolveu uma nova técnica para extrair proteína muscular que lhe permitiu isolar quantidades substanciais de actina relativamente pura , publicada em 1942. O método de Straub é essencialmente o mesmo usado em laboratórios hoje. Como a proteína de Straub era necessária para ativar a coagulação da miosina, ela foi chamada de actina . Percebendo que as preparações de miosina coagulante de Banga também continham actina, Szent-Györgyi chamou a mistura de ambas as proteínas de actomiosina .
As hostilidades da Segunda Guerra Mundial impediram que Szent-Gyorgyi publicasse o trabalho de seu laboratório em revistas científicas ocidentais . A Actin, portanto, só se tornou conhecida no Ocidente em 1945, quando seu artigo foi publicado como um suplemento da Acta Physiologica Scandinavica . Straub continuou a trabalhar com a actina e, em 1950, relatou que a actina contém ATP ligado e que, durante a polimerização da proteína em microfilamentos , o nucleotídeo é hidrolisado em ADP e fosfato inorgânico (que permanecem ligados ao microfilamento). Straub sugeriu que a transformação da actina ligada ao ATP em actina ligada ao ADP desempenhou um papel na contração muscular. Na verdade, isso é verdade apenas no músculo liso e não foi sustentado por meio de experimentação até 2001.
O sequenciamento de aminoácidos da actina foi concluído por M. Elzinga e colaboradores em 1973. A estrutura cristalina da G-actina foi resolvida em 1990 por Kabsch e colegas. No mesmo ano, um modelo para F-actina foi proposto por Holmes e colegas após experimentos usando co-cristalização com diferentes proteínas. O procedimento de cocristalização com diferentes proteínas foi utilizado repetidamente nos anos seguintes, até que em 2001 a proteína isolada cristalizou junto com o ADP. No entanto, ainda não existe uma estrutura de raios-X de alta resolução da F-actina. A cristalização da F-actina foi possível devido ao uso de um conjugado de rodamina que impede a polimerização pelo bloqueio do aminoácido cys-374 . Christine Oriol-Audit morreu no mesmo ano em que a actina foi cristalizada pela primeira vez, mas ela foi a pesquisadora que em 1977 cristalizou pela primeira vez a actina na ausência de Proteínas de Ligação à Actina (ABPs). No entanto, os cristais resultantes eram muito pequenos para a tecnologia disponível na época.
Embora nenhum modelo de alta resolução da forma filamentosa da actina exista atualmente, em 2008 a equipe de Sawaya foi capaz de produzir um modelo mais exato de sua estrutura com base em múltiplos cristais de dímeros de actina que se ligam em diferentes lugares. Este modelo foi posteriormente refinado por Sawaya e Lorenz. Outras abordagens, como o uso de microscopia crioeletrônica e radiação síncrotron , permitiram recentemente aumentar a resolução e melhor compreender a natureza das interações e mudanças conformacionais implicadas na formação dos filamentos de actina.
Estrutura
A sequência de aminoácidos da actina é uma das proteínas mais conservadas , pois mudou pouco ao longo da evolução , diferindo em não mais do que 20% em espécies tão diversas como algas e humanos . Portanto, considera-se que possui uma estrutura otimizada . Possui duas características distintivas: é uma enzima que hidroliza lentamente o ATP , a "moeda universal de energia" dos processos biológicos. No entanto, o ATP é necessário para manter sua integridade estrutural. Sua estrutura eficiente é formada por um processo de dobramento quase único . Além disso, é capaz de realizar mais interações do que qualquer outra proteína, o que lhe permite realizar uma ampla variedade de funções do que outras proteínas em quase todos os níveis da vida celular. A miosina é um exemplo de proteína que se liga à actina. Outro exemplo é a vilina , que pode tecer a actina em feixes ou cortar os filamentos dependendo da concentração de cátions de cálcio no meio circundante.
A actina é uma das proteínas mais abundantes em eucariotos , onde é encontrada em todo o citoplasma. Na verdade, nas fibras musculares , compreende 20% da proteína celular total em peso e entre 1% e 5% nas outras células. No entanto, não existe apenas um tipo de actina; os genes que codificam para a actina são definidos como uma família de genes (uma família que nas plantas contém mais de 60 elementos, incluindo genes e pseudogenes e em humanos mais de 30 elementos). Isso significa que a informação genética de cada indivíduo contém instruções que geram variantes de actina (chamadas isoformas ) que possuem funções ligeiramente diferentes. Isso, por sua vez, significa que os organismos eucarióticos expressam diferentes genes que dão origem a: α-actina, que é encontrada em estruturas contráteis; β-actina, encontrada na borda em expansão das células que usam a projeção de suas estruturas celulares como meio de mobilidade; e γ-actina, que é encontrada nos filamentos das fibras de estresse . Além das semelhanças que existem entre as isoformas de um organismo, há também uma conservação evolutiva na estrutura e função, mesmo entre organismos contidos em diferentes domínios eucarióticos . Em bactérias , foi identificado o homólogo de actina MreB , que é uma proteína capaz de polimerizar em microfilamentos; e em archaea o homólogo Ta0583 é ainda mais semelhante às actins eucarióticas.
A actina celular tem duas formas: glóbulos monoméricos chamados G-actina e filamentos poliméricos chamados F-actina (isto é, como filamentos compostos de muitos monômeros G-actina). A F-actina também pode ser descrita como um microfilamento. Dois fios paralelos de F-actina devem girar 166 graus para ficarem corretamente um sobre o outro. Isso cria a estrutura de dupla hélice dos microfilamentos encontrados no citoesqueleto. Os microfilamentos medem aproximadamente 7 nm de diâmetro com a hélice se repetindo a cada 37 nm. Cada molécula de actina está ligada a uma molécula de trifosfato de adenosina (ATP) ou difosfato de adenosina (ADP) que está associada a um cátion Mg 2+ . As formas mais comumente encontradas de actina, em comparação com todas as combinações possíveis, são ATP-G-Actina e ADP-F-actina.
G-actina
As imagens do microscópio eletrônico de varredura indicam que a G-actina tem uma estrutura globular; entretanto, a cristalografia de raios X mostra que cada um desses glóbulos consiste em dois lobos separados por uma fenda. Esta estrutura representa a “dobra de ATPase”, que é um centro de catálise enzimática que liga ATP e Mg 2+ e hidrolisa o anterior em ADP mais fosfato . Esta dobra é um motivo estrutural conservado que também é encontrado em outras proteínas que interagem com nucleotídeos trifosfato , como hexoquinase (uma enzima usada no metabolismo energético ) ou em proteínas Hsp70 (uma família de proteínas que desempenha um papel importante no enovelamento de proteínas). A G-actina só é funcional quando contém ADP ou ATP em sua fenda, mas a forma que está ligada ao ATP predomina nas células quando a actina está presente em seu estado livre.

O modelo de cristalografia de raios-X de actina que foi produzido por Kabsch a partir do tecido muscular estriado de coelhos é o mais comumente usado em estudos estruturais, pois foi o primeiro a ser purificado . A G-actina cristalizada por Kabsch tem aproximadamente 67 x 40 x 37 Å de tamanho, tem uma massa molecular de 41.785 Da e um ponto isoelétrico estimado de 4,8. Sua carga líquida em pH = 7 é -7.
- Estrutura primária
Elzinga e colaboradores determinaram pela primeira vez a sequência completa do peptídeo para este tipo de actina em 1973, com trabalhos posteriores do mesmo autor adicionando mais detalhes ao modelo. Ele contém 374 resíduos de aminoácidos . Seu terminal N é altamente ácido e começa com um aspartato acetilado em seu grupo amino. Enquanto seu terminal C é alcalino e é formado por uma fenilalanina precedida por uma cisteína , que tem um grau de importância funcional. Ambos os extremos estão próximos dentro do subdomínio I. Uma N τ -metil - histidina anômala está localizada na posição 73.
- Estrutura terciária - domínios
A estrutura terciária é formada por dois domínios conhecidos como o grande e o pequeno, que são separados por uma fenda centrada em torno da localização da ligação com ATP - ADP + P i . Abaixo deste existe um entalhe mais profundo denominado “ranhura”. No estado nativo , apesar de seus nomes, ambos têm uma profundidade comparável.
A convenção normal em estudos topológicos significa que uma proteína é mostrada com o maior domínio no lado esquerdo e o menor domínio no lado direito. Nesta posição, o domínio menor é, por sua vez, dividido em dois: subdomínio I (posição inferior, resíduos 1-32, 70-144 e 338-374) e subdomínio II (posição superior, resíduos 33-69). O domínio maior também é dividido em dois: subdomínio III (inferior, resíduos 145-180 e 270-337) e subdomínio IV (superior, resíduos 181-269). As áreas expostas dos subdomínios I e III são denominadas extremidades “farpadas”, enquanto as áreas expostas dos domínios II e IV são denominadas extremidades “pontiagudas”. Esta nomenclatura refere-se ao fato de que, devido à pequena massa do subdomínio A actina II é polar, a importância disso será discutida a seguir na discussão sobre a dinâmica de montagem.Alguns autores chamam os subdomínios Ia, Ib, IIa e IIb, respectivamente.
- Outras estruturas importantes
A estrutura supersecundária mais notável é uma folha beta de cinco cadeias que é composta por um β-meandro e uma unidade β-α-β no sentido horário. Está presente em ambos os domínios, sugerindo que a proteína surgiu da duplicação do gene.
- O local de ligação do nucleotídeo de adenosina está localizado entre duas estruturas em forma de grampo beta pertencentes aos domínios I e III. Os resíduos que estão envolvidos são Asp11-Lys18 e Asp154-His161, respectivamente.
- O sítio de ligação do cátion divalente está localizado logo abaixo daquele do nucleotídeo de adenosina. In vivo é mais frequentemente formado por Mg 2+ ou Ca 2+, enquanto in vitro é formado por uma estrutura quelante composta por Lys18 e dois oxigênios dos fosfatos α e β do nucleotídeo . Este cálcio é coordenado com seis moléculas de água que são retidas pelos aminoácidos Asp11 , Asp154 e Gln137 . Formam um complexo com o nucleotídeo que restringe os movimentos da região chamada "dobradiça", localizada entre os resíduos 137 e 144. Isso mantém a forma nativa da proteína até que sua retirada desnatura o monômero de actina. Essa região também é importante porque determina se a fenda da proteína está na conformação "aberta" ou "fechada".
- É altamente provável que existam pelo menos três outros centros com menor afinidade (intermediário) e ainda outros com baixa afinidade para cátions divalentes. Tem sido sugerido que esses centros podem desempenhar um papel na polimerização da actina, agindo durante a fase de ativação.
- Há uma estrutura em subdomínio 2 que é chamado de “D-circuito” porque se liga com ADNase I , que está localizado entre as His40 e Gly48 resíduos. Tem a aparência de um elemento desordenado na maioria dos cristais, mas parece uma folha β quando é complexado com DNase I. Foi proposto que o evento chave na polimerização é provavelmente a propagação de uma mudança conformacional do centro da ligação com o nucleotídeo a este domínio, que muda de um loop para uma espiral. No entanto, essa hipótese foi refutada por outros estudos.
F-actina

A descrição clássica de F-actina estados que tem uma estrutura filamentosa, que pode ser considerado como sendo uma única cadeia levógiro hélice com uma rotação de 166 ° em torno do eixo helicoidal e uma translação axial de 27,5 Â , ou uma única cadeia dextrógira hélice com um espaçamento cruzado de 350–380 Å, com cada actina cercada por mais quatro. A simetria do polímero de actina em 2,17 subunidades por volta de uma hélice é incompatível com a formação de cristais , que só é possível com uma simetria de exatamente 2, 3, 4 ou 6 subunidades por volta. Portanto, modelos devem ser construídos para explicar essas anomalias usando dados de microscopia eletrônica , microscopia crioeletrônica , cristalização de dímeros em diferentes posições e difração de raios-X . Vale ressaltar que não é correto falar em “estrutura” para uma molécula tão dinâmica quanto o filamento de actina. Na realidade, falamos de estados estruturais distintos, nestes a medição da translação axial permanece constante em 27,5 Å enquanto os dados de rotação da subunidade mostram uma variabilidade considerável, com deslocamentos de até 10% de sua posição ótima comumente vista. Algumas proteínas, como a cofilina, parecem aumentar o ângulo de giro, mas, novamente, isso pode ser interpretado como o estabelecimento de diferentes estados estruturais. Isso pode ser importante no processo de polimerização.
Há menos concordância em relação às medições do raio de giro e espessura do filamento: enquanto os primeiros modelos atribuíram um comprimento de 25 Å, os dados atuais de difração de raios-X, apoiados por microscopia crioeletrônica, sugerem um comprimento de 23,7 Å. Esses estudos mostraram os pontos de contato precisos entre os monômeros. Alguns são formados com unidades da mesma cadeia, entre a extremidade "farpada" de um monômero e a extremidade "pontiaguda" do próximo. Enquanto os monômeros em cadeias adjacentes fazem contato lateral por meio de projeções do subdomínio IV, com as projeções mais importantes sendo aquelas formadas pelo terminal C e a ligação hidrofóbica formada por três corpos envolvendo os resíduos 39–42, 201–203 e 286. O modelo sugere que um filamento é formado por monômeros em uma formação de "folha", na qual os subdomínios giram em torno de si mesmos, esta forma também é encontrada no homólogo de actina bacteriana MreB .
O polímero F-actina é considerado ter polaridade estrutural devido ao fato de que todas as subunidades do microfilamento apontam para a mesma extremidade. Isso dá origem a uma convenção de nomenclatura: a extremidade que possui uma subunidade de actina que tem seu local de ligação de ATP exposto é chamada de "extremidade (-)", enquanto a extremidade oposta, onde a fenda é direcionada a um monômero adjacente diferente, é chamada de " (+) fim ". Os termos "pontiagudo" e "farpado" que se referem às duas extremidades dos microfilamentos derivam de sua aparência à microscopia eletrônica de transmissão quando as amostras são examinadas seguindo uma técnica de preparação chamada "decoração". Este método consiste na adição de fragmentos de miosina S1 ao tecido que foi fixado com ácido tânico . Essa miosina forma ligações polares com monômeros de actina, dando origem a uma configuração que se parece com flechas com penas ao longo de sua haste, onde a haste é a actina e as penas são a miosina. Seguindo essa lógica, a extremidade do microfilamento que não possui nenhuma miosina saliente é chamada de ponta da seta (extremidade -) e a outra extremidade é chamada de extremidade farpada (extremidade +). Um fragmento S1 é composto pelos domínios da cabeça e pescoço da miosina II . Em condições fisiológicas, a G-actina (a forma monomérica ) é transformada em F-actina (a forma polimérica ) pelo ATP, onde o papel do ATP é essencial.
O filamento helicoidal F-actina encontrado nos músculos também contém uma molécula de tropomiosina , que é uma proteína de 40 nanômetros que envolve a hélice F-actina. Durante a fase de repouso, a tropomiosina cobre os locais ativos da actina, de modo que a interação actina-miosina não pode ocorrer e produzir contração muscular. Há outras moléculas de proteína ligadas ao fio de tropomiosina, estas são as troponinas que têm três polímeros: a troponina I , a troponina T e a troponina C .
Dobrando

A actina pode adquirir espontaneamente grande parte de sua estrutura terciária . No entanto, a maneira como ele adquire sua forma totalmente funcional de sua forma nativa recém- sintetizada é especial e quase única na química de proteínas. A razão para esta rota especial pode ser a necessidade de evitar a presença de monômeros de actina incorretamente dobrados, que podem ser tóxicos, pois podem atuar como terminadores de polimerização ineficientes. No entanto, é fundamental para estabelecer a estabilidade do citoesqueleto e, além disso, é um processo essencial para a coordenação do ciclo celular .
A CCT é necessária para garantir que a dobra ocorra corretamente. CCT é uma chaperonina do grupo II, um grande complexo de proteínas que auxilia no enovelamento de outras proteínas. A CCT é formada por um anel duplo de oito subunidades diferentes (hetero-octamérica) e difere das chaperoninas do grupo I, como GroEL , que é encontrada em eubactérias e organelas eucarióticas, pois não requer uma co-chaperona para atuar como uma tampa sobre a cavidade catalítica central . Os substratos ligam-se ao CCT por meio de domínios específicos. Inicialmente, pensou-se que se ligava apenas à actina e à tubulina , embora estudos recentes de imunoprecipitação tenham mostrado que ela interage com um grande número de polipeptídeos , que possivelmente funcionam como substratos . Ele age por meio de mudanças conformacionais dependentes de ATP que às vezes requerem várias rodadas de liberação e catálise para completar uma reação.
Para completar com sucesso seu enovelamento, a actina e a tubulina precisam interagir com outra proteína chamada prefoldina , que é um complexo heterohexamérico (formado por seis subunidades distintas), em uma interação tão específica que as moléculas coevoluem . A actina forma um complexo com a prefoldina enquanto ela ainda está sendo formada, quando ela tem aproximadamente 145 aminoácidos de comprimento, especificamente aqueles no N-terminal.
Diferentes subunidades de reconhecimento são usadas para actina ou tubulina, embora haja alguma sobreposição. Na actina, as subunidades que se ligam à prefoldina são provavelmente PFD3 e PFD4, que se ligam em dois locais, um entre os resíduos 60–79 e o outro entre os resíduos 170–198. A actina é reconhecida, carregada e fornecida à chaperonina citosólica (CCT) em uma conformação aberta pela extremidade interna dos "tentáculos" da pré-moldina (veja a imagem e nota). O contato quando a actina é fornecida é tão breve que um complexo terciário não é formado, liberando imediatamente a prefoldina.

O CCT, então, causa o dobramento sequencial da actina, formando ligações com suas subunidades, em vez de simplesmente encerrá-la em sua cavidade. É por isso que possui áreas de reconhecimento específicas em seu domínio β apical. A primeira etapa do dobramento consiste no reconhecimento dos resíduos 245–249. Em seguida, outros determinantes estabelecem contato. Tanto a actina quanto a tubulina se ligam ao CCT em conformações abertas na ausência de ATP. No caso da actina, duas subunidades são ligadas durante cada mudança conformacional, enquanto para a tubulina a ligação ocorre com quatro subunidades. A actina tem sequências de ligação específicas, que interagem com as subunidades δ e β-CCT ou com δ-CCT e ε-CCT. Depois que o AMP-PNP é ligado ao CCT, os substratos se movem dentro da cavidade da chaperonina. Parece também que, no caso da actina, a proteína CAP é necessária como um possível cofator nos estados finais de dobramento da actina.
A maneira exata pela qual esse processo é regulado ainda não está totalmente esclarecida, mas sabe-se que a proteína PhLP3 (proteína semelhante à fosducina ) inibe sua atividade por meio da formação de um complexo terciário.
Mecanismo catalítico da ATPase
A actina é uma ATPase , o que significa que é uma enzima que hidrolisa o ATP. Este grupo de enzimas é caracterizado por suas taxas de reação lentas. Sabe-se que essa ATPase é “ativa”, ou seja, sua velocidade aumenta cerca de 40.000 vezes quando a actina faz parte de um filamento. Um valor de referência para essa taxa de hidrólise em condições ideais é em torno de 0,3 s -1 . Então, o P i permanece ligado à actina ao lado do ADP por um longo tempo, até que seja cooperativamente liberado do interior do filamento.
Os detalhes moleculares exatos do mecanismo catalítico ainda não são totalmente compreendidos. Embora haja muito debate sobre o assunto, parece certo que uma conformação "fechada" é necessária para a hidrólise do ATP, e acredita-se que os resíduos que estão envolvidos no processo se movam para a distância adequada. O ácido glutâmico Glu137 é um dos resíduos-chave, que está localizado no subdomínio 1. Sua função é ligar a molécula de água que produz um ataque nucleofílico na ligação γ-fosfato do ATP , enquanto o nucleotídeo está fortemente ligado aos subdomínios 3 e 4 A lentidão do processo catalítico deve-se à grande distância e posição enviesada da molécula de água em relação ao reagente. É altamente provável que a mudança conformacional produzida pela rotação dos domínios entre as formas G e F da actina mova o Glu137 para mais perto, permitindo sua hidrólise. Este modelo sugere que a polimerização e a função da ATPase seriam desacopladas imediatamente. A transformação "aberta" para "fechada" entre as formas G e F e suas implicações no movimento relativo de vários resíduos-chave e na formação de fios d'água foram caracterizadas em dinâmica molecular e simulações QM / MM .
Genética

A actina tem sido uma das proteínas mais conservadas ao longo da evolução porque interage com um grande número de outras proteínas. Possui 80,2% de conservação de sequência no nível do gene entre Homo sapiens e Saccharomyces cerevisiae (uma espécie de levedura) e 95% de conservação da estrutura primária do produto proteico.
Embora a maioria das leveduras tenha apenas um único gene de actina, eucariotos superiores , em geral, expressam várias isoformas de actina codificadas por uma família de genes relacionados. Os mamíferos têm pelo menos seis isoformas de actina codificadas por genes separados, que são divididos em três classes (alfa, beta e gama) de acordo com seus pontos isoelétricos . Em geral, as alfa actinas são encontradas no músculo (α-esquelético, α-liso aórtico, α-cardíaco), enquanto as isoformas beta e gama são proeminentes em células não musculares (β-citoplasmático, γ1-citoplasmático, γ2-liso entérico) . Embora as sequências de aminoácidos e as propriedades in vitro das isoformas sejam altamente semelhantes, essas isoformas não podem substituir completamente uma pela outra in vivo .
O gene típico da actina tem uma UTR 5 'de aproximadamente 100 nucleotídeos , uma região traduzida de 1200 nucleotídeos e uma UTR 3' de 200 nucleotídeos . A maioria dos genes da actina é interrompida por íntrons , com até seis íntrons em qualquer uma das 19 localizações bem caracterizadas. A alta conservação da família torna a actina o modelo preferido para estudos comparando os modelos íntrons-inicial e íntrons-tardio da evolução do íntron.
Todos os procariotos não esféricos parecem possuir genes como MreB , que codificam homólogos de actina; esses genes são necessários para que a forma da célula seja mantida. O gene ParM derivado do plasmídeo codifica uma proteína semelhante à actina cuja forma polimerizada é dinamicamente instável e parece particionar o DNA do plasmídeo em suas células filhas durante a divisão celular por um mecanismo análogo ao empregado pelos microtúbulos na mitose eucariótica . A actina é encontrada em retículos endoplasmáticos lisos e rugosos.
Dinâmica de montagem
Nucleação e polimerização

Fatores de nucleação são necessários para estimular a polimerização da actina. Um desses fatores de nucleação é o complexo Arp2 / 3 , que imita um dímero G-actina a fim de estimular a nucleação (ou formação do primeiro trímero) da G-actina monomérica. O complexo Arp2 / 3 liga-se aos filamentos de actina a 70 graus para formar novos ramos de actina a partir dos filamentos de actina existentes. A nucleação mediada por Arp2 / 3 é necessária para a migração celular direcionada. Além disso, os próprios filamentos de actina se ligam ao ATP, e a hidrólise desse ATP estimula a desestabilização do polímero.
O crescimento dos filamentos de actina pode ser regulado pela timosina e profilina . A timosina se liga à G-actina para tamponar o processo de polimerização, enquanto a profilina se liga à G-actina para trocar ADP por ATP , promovendo a adição monomérica ao farpado, além da extremidade dos filamentos de F-actina.
A F-actina é forte e dinâmica. Ao contrário de outros polímeros , como o DNA , cujos elementos constituintes são unidos por ligações covalentes , os monômeros dos filamentos de actina são montados por ligações mais fracas. As ligações laterais com monômeros vizinhos resolvem essa anomalia, que em teoria deveria enfraquecer a estrutura, pois podem ser rompidas por agitação térmica. Além disso, as ligações fracas oferecem a vantagem de que as extremidades dos filamentos podem facilmente liberar ou incorporar monômeros. Isso significa que os filamentos podem ser remodelados rapidamente e podem alterar a estrutura celular em resposta a um estímulo ambiental. Que, juntamente com o mecanismo bioquímico pelo qual é provocado, é conhecido como "dinâmica de montagem".
- Estudos in vitro
Os estudos que enfocam o acúmulo e a perda de subunidades por microfilamentos são realizados in vitro (ou seja, em laboratório e não em sistemas celulares), pois a polimerização da actina resultante dá origem à mesma F-actina produzida in vivo . O processo in vivo é controlado por uma infinidade de proteínas para torná-lo responsivo às demandas celulares, o que dificulta a observação de suas condições básicas.
A produção in vitro ocorre de forma sequencial: primeiro, ocorre a "fase de ativação", quando ocorre a ligação e a troca de cátions divalentes em locais específicos da G-actina, que está ligada ao ATP. Isso produz uma mudança conformacional, às vezes chamada de monômero G * -actina ou F-actina, pois é muito semelhante às unidades localizadas no filamento. Isso o prepara para a "fase de nucleação", na qual a G-actina dá origem a pequenos fragmentos instáveis de F-actina que são capazes de polimerizar. Dímeros e trímeros instáveis são inicialmente formados. A "fase de alongamento" começa quando há um número suficientemente grande desses polímeros curtos. Nesta fase, o filamento se forma e cresce rapidamente por meio da adição reversível de novos monômeros em ambos os extremos. Finalmente, um equilíbrio estacionário é alcançado onde os monômeros de G-actina são trocados em ambas as extremidades do microfilamento sem qualquer alteração em seu comprimento total. Nesta última fase a "concentração crítica C c " é definida como a razão entre a constante de montagem e a constante de dissociação para G-actina, onde a dinâmica de adição e eliminação de dímeros e trímeros não produz uma mudança no comprimento do microfilamento . Em condições in vitro , C c é 0,1 μM, o que significa que em valores mais altos ocorre a polimerização e em valores mais baixos ocorre a despolimerização.
- Papel da hidrólise de ATP
Conforme indicado acima, embora a actina hidrolise ATP, tudo aponta para o fato de que o ATP não é necessário para a actina ser montada, uma vez que, por um lado, a hidrólise ocorre principalmente dentro do filamento, e por outro lado o ADP também poderia instigar a polimerização. Isso levanta a questão de compreender qual processo termodinamicamente desfavorável requer tal prodigioso dispêndio de energia . O ciclo da actina, que acopla a hidrólise de ATP à polimerização de actina, consiste na adição preferencial de monômeros G-actina-ATP à extremidade farpada de um filamento e na desmontagem simultânea de monômeros F-actina-ADP na extremidade pontiaguda onde o ADP é subsequentemente mudou para ATP, fechando assim o ciclo. Este aspecto da formação do filamento de actina é conhecido como “treadmilling”.
O ATP é hidrolisado de forma relativamente rápida logo após a adição de um monômero de G-actina ao filamento. Existem duas hipóteses sobre como isso ocorre; o estocástico , que sugere que a hidrólise ocorre aleatoriamente de uma maneira que é de alguma forma influenciada pelas moléculas vizinhas; e a vetorial, que sugere que a hidrólise só ocorre adjacente a outras moléculas cujo ATP já foi hidrolisado. Em qualquer um dos casos, o P i resultante não é liberado; permanece por algum tempo ligado não covalentemente ao ADP da actina. Desta forma, existem três espécies de actina em um filamento: ATP-Actina, ADP + P i -Actina e ADP-Actina. A quantidade de cada uma dessas espécies presentes em um filamento depende de seu comprimento e estado: conforme o alongamento começa, o filamento tem uma quantidade aproximadamente igual de monômeros de actina ligados com ATP e ADP + P i e uma pequena quantidade de ADP-Actina no (-) fim. Quando o estado estacionário é alcançado, a situação se inverte, com ADP presente ao longo da maior parte do filamento e apenas a área mais próxima da extremidade (+) contendo ADP + P i e com ATP presente apenas na ponta.
Se compararmos os filamentos que contêm apenas ADP-Actina com aqueles que incluem ATP, no primeiro as constantes críticas são semelhantes em ambas as extremidades, enquanto C c para os outros dois nucleotídeos é diferente: Na extremidade (+) Cc + = 0,1 μM, enquanto no final (-) Cc - = 0,8 μM, o que dá origem às seguintes situações:
- Para concentrações de G-actina-ATP inferiores a Cc +, não ocorre alongamento do filamento.
- Para concentrações de G-actina-ATP menores que Cc - mas maiores que Cc +, o alongamento ocorre na extremidade (+).
- Para concentrações de G-actina-ATP maiores que Cc - o microfilamento cresce em ambas as extremidades.
Portanto, é possível deduzir que a energia produzida pela hidrólise é utilizada para criar um verdadeiro “estado estacionário”, ou seja, um fluxo, ao invés de um equilíbrio simples, dinâmico, polar e aderido ao filamento. Isso justifica o gasto de energia, pois promove funções biológicas essenciais. Além disso, a configuração dos diferentes tipos de monômeros é detectada por proteínas de ligação à actina, que também controlam esse dinamismo, conforme será descrito na seção seguinte.
A formação de microfilamentos por esteira foi considerada atípica em estereocílios . Nesse caso, o controle do tamanho da estrutura é totalmente apical e é controlado de alguma forma pela expressão gênica, ou seja, pela quantidade total de monômero proteico sintetizado em determinado momento.
Proteínas associadas

O citoesqueleto de actina in vivo não é composto exclusivamente de actina, outras proteínas são necessárias para sua formação, continuidade e função. Essas proteínas são chamadas de proteínas de ligação à actina (ABP) e estão envolvidas na polimerização, despolimerização, estabilidade, organização em feixes ou redes, fragmentação e destruição da actina. A diversidade dessas proteínas é tal que acredita-se que a actina seja a proteína que participa do maior número de interações proteína-proteína . Por exemplo, existem elementos sequestrantes de G-actina que impedem sua incorporação em microfilamentos. Existem também proteínas que estimulam sua polimerização ou que complexificam as redes de síntese.
- A timosina β-4 é uma proteína de 5 kDa que pode se ligar ao G-actina-ATP em uma estequiometria de 1: 1 ; o que significa que uma unidade de timosina β-4 se liga a uma unidade de G-actina. Seu papel é impedir a incorporação dos monômeros no polímero em crescimento.
- A profilina , é uma proteína citosólica com peso molecular de 15 kDa, que também se liga à G-actina-ATP ou -ADP com estequiometria de 1: 1, mas tem uma função diferente, pois facilita a substituição de nucleotídeos de ADP por ATP . Também está implicado em outras funções celulares, como a ligação de repetições de prolina em outras proteínas ou de lipídios que atuam como mensageiros secundários .

Outras proteínas que se ligam à actina regulam o comprimento dos microfilamentos, cortando-os, o que dá origem a novas extremidades ativas para a polimerização. Por exemplo, se um microfilamento com duas pontas for cortado duas vezes, haverá três novos microfilamentos com seis pontas. Esta nova situação favorece a dinâmica de montagem e desmontagem. As mais notáveis dessas proteínas são a gelsolina e a cofilina . Essas proteínas primeiro atingem um corte ligando-se a um monômero de actina localizado no polímero; então, alteram a conformação do monômero de actina enquanto permanecem ligadas à extremidade (+) recém-gerada. Isso tem o efeito de impedir a adição ou troca de novas subunidades de G-actina. A despolimerização é encorajada porque as extremidades (-) não estão ligadas a nenhuma outra molécula.
Outras proteínas que se ligam à actina cobrem as extremidades da F-actina para estabilizá-las, mas são incapazes de quebrá-las. Exemplos desse tipo de proteína são CapZ , que se liga às extremidades (+) dependendo dos níveis de Ca 2+ / calmodulina de uma célula . Esses níveis dependem dos sinais internos e externos da célula e estão envolvidos na regulação de suas funções biológicas). Outro exemplo é a tropomodulina (que se liga à extremidade (-)). A tropomodulina atua basicamente para estabilizar a F-actina presente nas miofibrilas presentes nos sarcômeros musculares , estruturas caracterizadas por sua grande estabilidade.

O complexo Arp2 / 3 é amplamente encontrado em todos os organismos eucarióticos . É composto por sete subunidades, algumas das quais possuem uma topologia claramente relacionada à sua função biológica: duas das subunidades, ARP2 e ARP3, têm uma estrutura semelhante à dos monômeros de actina. Esta homologia permite que ambas as unidades atuem como agentes de nucleação na polimerização de G-actina e F-actina. Este complexo também é necessário em processos mais complicados, como no estabelecimento de estruturas dendríticas e também na anastomose (a reconexão de duas estruturas ramificadas que haviam sido previamente unidas, como nos vasos sanguíneos).
Inibidores químicos

Há uma série de toxinas que interferem na dinâmica da actina, seja impedindo-a de polimerizar ( latrunculina e citocalasina D ) ou estabilizando-a ( faloidina ):
- Latrunculin é uma toxina produzida por esponjas . Ele se liga à G-actina impedindo-a de se ligar aos microfilamentos.
- A citocalasina D, é um alcalóide produzido por fungos , que se liga à extremidade (+) da F-actina impedindo a adição de novos monômeros. Foi descoberto que a citocalasina D interrompe a dinâmica da actina, ativando a proteína p53 em animais.
- Phalloidin, é uma toxina que foi isolada do cogumelo Amanita phalloides . Ele se liga à interface entre os monômeros de actina adjacentes no polímero F-actina, evitando sua despolimerização.
Funções e localização
Os filamentos das formas de actina ('F-actina' ou microfilamentos ) são elementos essenciais do citoesqueleto eucariótico , capazes de sofrer uma dinâmica de polimerização e despolimerização muito rápida. Na maioria das células, os filamentos de actina formam redes em larga escala que são essenciais para muitas funções-chave nas células:
- Vários tipos de redes de actina (feitas de filamentos de actina) fornecem suporte mecânico às células e fornecem rotas de tráfego através do citoplasma para auxiliar na transdução do sinal.
- A rápida montagem e desmontagem da rede de actina permite que as células migrem ( Migração celular ).
- Nas células musculares dos metazoários , é o arcabouço no qual as proteínas da miosina geram força para suportar a contração muscular.
- Em células não musculares, ser uma via para o transporte de carga de miosinas (miosinas não convencionais) como a miosina V e VI. As miosinas não convencionais usam a hidrólise de ATP para transportar cargas, como vesículas e organelas, de uma forma direcionada muito mais rápida do que a difusão. A miosina V caminha em direção à extremidade farpada dos filamentos de actina, enquanto a miosina VI caminha em direção à extremidade pontiaguda. A maioria dos filamentos de actina é organizada com a extremidade farpada em direção à membrana celular e a extremidade pontiaguda em direção ao interior da célula. Este arranjo permite que a miosina V seja um motor eficaz para a exportação de cargas e a miosina VI seja um motor eficaz para importação.
A proteína actina é encontrada no citoplasma e no núcleo da célula . Sua localização é regulada pelas vias de transdução de sinal da membrana celular que integram os estímulos que uma célula recebe estimulando a reestruturação das redes de actina em resposta. Em Dictyostelium , a fosfolipase D foi encontrada para intervir nas vias de fosfato de inositol . Os filamentos de actina são particularmente estáveis e abundantes nas fibras musculares . Dentro do sarcômero (a unidade morfológica e fisiológica básica das fibras musculares), a actina está presente nas bandas I e A; a miosina também está presente neste último.
Citoesqueleto

Os microfilamentos estão envolvidos no movimento de todas as células móveis, incluindo os tipos não musculares, e as drogas que interrompem a organização da F-actina (como as citocalasinas ) afetam a atividade dessas células. A actina compreende 2% da quantidade total de proteínas nos hepatócitos , 10% nos fibroblastos , 15% nas amebas e até 50–80% nas plaquetas ativadas . Existem vários tipos diferentes de actina com estruturas e funções ligeiramente diferentes. Isso significa que a α-actina é encontrada exclusivamente nas fibras musculares , enquanto os tipos β e γ são encontrados em outras células. Além disso, como os últimos tipos apresentam uma alta taxa de rotatividade, a maioria deles se encontra fora de estruturas permanentes. Isso significa que os microfilamentos encontrados em células diferentes das células musculares estão presentes em três formas:
- Redes de microfilamentos - as células animais geralmente têm um córtex celular sob a membrana celular que contém um grande número de microfilamentos, o que impede a presença de organelas . Essa rede está conectada a várias células receptoras que transmitem sinais para o exterior de uma célula.

- Feixes de microfilamentos - Esses microfilamentos extremamente longos estão localizados em redes e, em associação com proteínas contráteis, como a miosina não muscular , estão envolvidos no movimento de substâncias em um nível intracelular.
- Anéis de actina periódicos - descobriu-se recentemente que uma estrutura periódica construída de anéis de actina com espaçamento uniforme existe especificamente em axônios (não em dendritos ). Nessa estrutura, os anéis de actina, juntamente com os tetrâmeros de espectrina que ligam os anéis de actina vizinhos, formam um citoesqueleto coeso que suporta a membrana do axônio. A periodicidade da estrutura também pode regular os canais de íons de sódio nos axônios.
Fermento
O citoesqueleto da actina é fundamental para os processos de endocitose , citocinese , determinação da polaridade celular e morfogênese em leveduras . Além de depender da actina, esses processos envolvem 20 ou 30 proteínas associadas, todas com alto grau de conservação evolutiva, junto com muitas moléculas sinalizadoras. Juntos, esses elementos permitem uma montagem modulada espacial e temporalmente que define a resposta de uma célula a estímulos internos e externos.
As leveduras contêm três elementos principais que estão associados à actina: remendos, cabos e anéis que, apesar de não estarem presentes por muito tempo, estão sujeitos a um equilíbrio dinâmico devido à polimerização e despolimerização contínuas. Eles possuem uma série de proteínas acessórias, incluindo ADF / cofilina, que tem um peso molecular de 16 kDa e é codificada por um único gene, denominado COF1 ; Aip1, um cofator de cofilina que promove a desmontagem de microfilamentos; Srv2 / CAP, um regulador de processo relacionado às proteínas adenilato ciclase ; uma profilina com um peso molecular de aproximadamente 14 kDa que está relacionada / associada a monômeros de actina; e twinfilin, uma proteína de 40 kDa envolvida na organização de manchas.
Plantas
Estudos do genoma de plantas revelaram a existência de isovariantes de proteínas dentro da família de genes da actina. Dentro de Arabidopsis thaliana , um dicotiledóneo usado como organismo modelo , existem dez tipos de actina, nove tipos de α-tubulinas, seis β-tubulinas, seis profilinas e dezenas de miosinas. Essa diversidade é explicada pela necessidade evolutiva de possuir variantes que diferem ligeiramente em sua expressão temporal e espacial. A maioria dessas proteínas foi expressa em conjunto no tecido analisado. Redes de actina são distribuídas por todo o citoplasma das células que foram cultivadas in vitro . Há uma concentração da rede ao redor do núcleo que se conecta por meio de raios ao córtex celular, essa rede é altamente dinâmica, com polimerização e despolimerização contínuas.

Embora a maioria das células vegetais tenha uma parede celular que define sua morfologia e impede seu movimento, seus microfilamentos podem gerar força suficiente para atingir uma série de atividades celulares, como as correntes citoplasmáticas geradas pelos microfilamentos e miosina. A actina também está envolvida no movimento das organelas e na morfogênese celular, que envolve a divisão celular , bem como o alongamento e a diferenciação da célula.
As proteínas mais notáveis associadas ao citoesqueleto de actina em plantas incluem: vilina , que pertence à mesma família da gelsolina / severina e é capaz de cortar microfilamentos e ligar monômeros de actina na presença de cátions de cálcio; fimbrina , que é capaz de reconhecer e unir monômeros de actina e que está envolvida na formação de redes (por um processo de regulação diferente daquele de animais e leveduras); forminas , que são capazes de atuar como um agente de nucleação de polimerização de F-actina; miosina , um motor molecular típico específico para eucariotos e que em Arabidopsis thaliana é codificado por 17 genes em duas classes distintas; CHUP1, que pode se ligar à actina e está implicado na distribuição espacial dos cloroplastos na célula; KAM1 / MUR3 que definem a morfologia do aparelho de Golgi , bem como a composição de xiloglucanos na parede celular; NtWLIM1, que facilita o surgimento de estruturas celulares de actina; e ERD10, que está envolvido na associação de organelas dentro de membranas e microfilamentos e que parece desempenhar um papel que está envolvido na reação de um organismo ao estresse .
Actina nuclear
A actina nuclear foi notada e descrita pela primeira vez em 1977 por Clark e Merriam. Os autores descrevem uma proteína presente na fração nuclear, obtida de oócitos de Xenopus laevis , que apresenta as mesmas características da actina do músculo esquelético. Desde aquela época, houve muitos relatórios científicos sobre a estrutura e funções da actina no núcleo (para revisão, consulte: Hofmann 2009.) O nível controlado de actina no núcleo, sua interação com proteínas de ligação à actina (ABP) e a presença de diferentes isoformas permite que a actina desempenhe um papel importante em muitos processos nucleares importantes.
Transporte de actina através da membrana nuclear
A sequência de actina não contém um sinal de localização nuclear. O pequeno tamanho da actina (cerca de 43 kDa) permite que ela entre no núcleo por difusão passiva. A actina, entretanto, se move rapidamente entre o citoplasma e o núcleo, o que indica a existência de transporte ativo. A importação de actina para o núcleo (provavelmente em um complexo com cofilina) é facilitada pela importação de proteína importina 9.
O baixo nível de actina no núcleo parece ser muito importante, porque a actina tem dois sinais de exportação nuclear (NES) em sua sequência. A actina microinjetada é rapidamente removida do núcleo para o citoplasma. A actina é exportada pelo menos de duas maneiras, por meio da exportação na 1 (EXP1) e da exportação na 6 (Exp6).
Modificações específicas, como a SUMOilação, permitem a retenção de actina nuclear. Foi demonstrado que uma mutação que impede a SUMOilação causa rápida exportação de beta actina do núcleo.
Com base nos resultados experimentais, um mecanismo geral de transporte de actina nuclear pode ser proposto:
- No citoplasma, a cofilina se liga aos monômeros de ADP-actina. Este complexo é importado ativamente para o núcleo.
- A maior concentração de ATP no núcleo (em comparação com o citoplasma) promove a troca de ADP para ATP no complexo actina-cofilina. Isso enfraquece a força de ligação dessas duas proteínas.
- O complexo cofilina-actina finalmente se dissocia após a fosforilação da cofilina pela quinase LIM nuclear.
- A actina é SUMOilada e nesta forma é retida dentro do núcleo.
- A actina pode formar complexos com a profilina e deixar o núcleo via exportina 6.
A organização da actina nuclear
A actina nuclear existe principalmente como um monômero, mas também pode formar oligômeros dinâmicos e polímeros curtos. A organização da actina nuclear varia em diferentes tipos de células. Por exemplo, em oócitos Xenopus (com maior nível de actina nuclear em comparação com células somáticas) a actina forma filamentos, que estabilizam a arquitetura do núcleo. Esses filamentos podem ser observados ao microscópio graças à coloração com faloidina conjugada com fluoróforo.
Em núcleos de células somáticas, no entanto, os filamentos de actina não podem ser observados usando esta técnica. O ensaio de inibição da DNase I, até agora o único teste que permite a quantificação da actina polimerizada diretamente em amostras biológicas, revelou que a actina nuclear endógena de fato ocorre principalmente na forma monomérica.
O nível de actina precisamente controlado no núcleo da célula, mais baixo do que no citoplasma, evita a formação de filamentos. A polimerização também é reduzida pelo acesso limitado aos monômeros de actina, que se ligam em complexos com ABPs, principalmente a cofilina.
Isoformas de actina no núcleo da célula
Pouca atenção é dada às isoformas de actina; no entanto, foi demonstrado que diferentes isoformas de actina estão presentes no núcleo da célula. As isoformas de actina, apesar de sua alta similaridade de sequência, apresentam diferentes propriedades bioquímicas, como cinética de polimerização e despolimerização. Eles também mostram localizações e funções diferentes.
O nível de isoformas de actina, tanto no citoplasma quanto no núcleo, pode mudar, por exemplo, em resposta à estimulação do crescimento celular ou interrupção da proliferação e atividade transcricional.
As preocupações com a pesquisa sobre a actina nuclear geralmente se concentram na isoforma beta. No entanto, o uso de anticorpos dirigidos contra diferentes isoformas de actina permite identificar não apenas a beta citoplasmática no núcleo da célula, mas também:
- gama actina nos núcleos celulares do melanoma humano,
- Actina do músculo esquelético alfa nos núcleos dos mioblastos de camundongo,
- gama-actina citoplasmática e também alfa-actina de músculo liso no núcleo do fibroblasto fetal de camundongo
A presença de diferentes isoformas de actina pode ter um efeito significativo em sua função em processos nucleares, especialmente porque o nível de isoformas individuais pode ser controlado de forma independente.
Funções de actina nuclear
As funções da actina no núcleo estão associadas à sua capacidade de polimerizar e interagir com vários ABPs e com elementos estruturais do núcleo. A actina nuclear está envolvida em:
- Arquitetura do núcleo - A interação da actina com alfa II-espectrina e outras proteínas são importantes para manter a forma adequada do núcleo.
- Transcrição - a actina está envolvida na reorganização da cromatina, na iniciação da transcrição e na interação com o complexo de transcrição. A actina participa da regulação da estrutura da cromatina, interagindo com a RNA polimerase I, II e III. Na transcrição da Pol I, a actina e a miosina ( MYO1C , que se liga ao DNA) atuam como um motor molecular . Para a transcrição de Pol II, a β-actina é necessária para a formação do complexo de pré-iniciação. Pol III contém β-actina como uma subunidade. A actina também pode ser um componente de complexos de remodelação da cromatina, bem como de partículas pré-mRNP (isto é, RNA mensageiro precursor agrupado em proteínas) e está envolvida na exportação nuclear de RNAs e proteínas.
- Regulação da atividade do gene - a actina se liga às regiões reguladoras de diferentes tipos de genes. A capacidade da actina de regular a atividade gênica é utilizada no método de reprogramação molecular, que permite que células diferenciadas retornem ao estado embrionário.
- Translocação do fragmento do cromossomo ativado da região da membrana para a eucromatina, onde a transcrição começa. Este movimento requer a interação de actina e miosina.
- Integração de diferentes compartimentos celulares . A actina é uma molécula que integra as vias de transdução do sinal citoplasmático e nuclear. Um exemplo é a ativação da transcrição em resposta à estimulação sérica de células in vitro .
- Resposta imune - a actina nuclear polimeriza após a estimulação do receptor de células T e é necessária para a expressão de citocinas e produção de anticorpos in vivo .
Devido à sua capacidade de sofrer alterações conformacionais e interação com muitas proteínas, a actina atua como um regulador da formação e da atividade de complexos protéicos, como o complexo transcricional.
Contração muscular

Esboço de uma contração muscular
Nas células musculares, as miofibrilas de actomiosina constituem grande parte do material citoplasmático. Essas miofibrilas são feitas de filamentos finos de actina (tipicamente cerca de 7 nm de diâmetro) e filamentos grossos da proteína motora miosina (tipicamente cerca de 15 nm de diâmetro). Essas miofibrilas usam energia derivada do ATP para criar movimentos de células, como a contração muscular . Usando a hidrólise do ATP para obter energia, as cabeças de miosina passam por um ciclo durante o qual se prendem a filamentos finos, exercem uma tensão e, a seguir, dependendo da carga, executam um golpe de força que faz com que os filamentos finos deslizem, encurtando o músculo.
Em feixes contráteis, a proteína alfa- actinina do agrupamento de actina separa cada filamento fino em ~ 35 nm. Esse aumento na distância permite que filamentos grossos se encaixem e interajam, permitindo deformação ou contração. Na deformação, uma extremidade da miosina está ligada à membrana plasmática , enquanto a outra extremidade "caminha" em direção à extremidade positiva do filamento de actina. Isso puxa a membrana para uma forma diferente em relação ao córtex celular . Para a contração, a molécula de miosina é geralmente ligada a dois filamentos separados e ambas as extremidades simultaneamente "caminham" em direção à extremidade positiva do filamento, deslizando os filamentos de actina mais próximos um do outro. Isso resulta no encurtamento ou contração do feixe de actina (mas não do filamento). Esse mecanismo é responsável pela contração muscular e citocinese , a divisão de uma célula em duas.
O papel da actina na contração muscular
O filamento helicoidal de F-actina encontrado nos músculos também contém uma molécula de tropomiosina , uma proteína de 40 nanômetros que envolve a hélice de F-actina. Durante a fase de repouso, a tropomiosina cobre os sítios ativos da actina, de modo que a interação actina-miosina não pode ocorrer e produzir contração muscular (a interação dá origem a um movimento entre as duas proteínas que, por se repetir muitas vezes, produz uma contração) . Há outras moléculas de proteína ligadas ao fio de tropomiosina, estes incluem as troponinas que têm três polímeros: a troponina I , a troponina T e a troponina C . A função reguladora da tropomiosina depende de sua interação com a troponina na presença de íons Ca 2+ .
Tanto a actina quanto a miosina estão envolvidas na contração e relaxamento muscular e constituem 90% da proteína muscular. O processo geral é iniciado por um sinal externo, tipicamente por meio de um potencial de ação estimulando o músculo, que contém células especializadas cujos interiores são ricos em filamentos de actina e miosina. O ciclo de contração-relaxamento compreende as seguintes etapas:
- Despolarização do sarcolema e transmissão de um potencial de ação através dos túbulos T .
- Abertura do retículo sarcoplasmático de Ca 2+ canais.
- Aumento das concentrações citosólicas de Ca 2+ e a interação desses cátions com a troponina causando uma alteração conformacional em sua estrutura . Isso, por sua vez, altera a estrutura da tropomiosina, que cobre o sítio ativo da actina, permitindo a formação de ligações cruzadas de miosina-actina (esta última presente como filamentos finos).
- O movimento das cabeças de miosina sobre os filamentos finos pode envolver ATP ou ser independente do ATP. O anterior mecanismo, mediado por ATPase actividade nas cabeças de miosina, faz com que o movimento dos filamentos de actina em relação ao Z-disco .
- Captura de Ca 2+ pelo retículo sarcoplasmático, causando uma nova mudança conformacional na tropomiosina que inibe a interação actina-miosina.
Outros processos biológicos
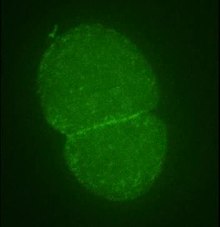
A imagem tradicional da função da actina relaciona-a à manutenção do citoesqueleto e, portanto, à organização e movimentação das organelas, bem como à determinação da forma de uma célula. No entanto, a actina tem um papel mais amplo na fisiologia das células eucarióticas, além de funções semelhantes em procariotos .
- Citocinese . A divisão celular em células animais e leveduras normalmente envolve a separação da célula-mãe em duas células-filhas por meio da constrição da circunferência central. Este processo envolve um anel constritor composto de actina, miosina, anilina e α-actinina . Na levedura de fissão Schizosaccharomyces pombe , a actina é formado activamente no anel constrição com a participação de Arp3 , o formin Cdc12, profilina , e WASp , juntamente com os microfilamentos de pré-formados. Uma vez que o anel foi construído, a estrutura é mantida por uma contínua montagem e desmontagem que, auxiliada pelo complexo Arp2 / 3 e pelas formas, é a chave para um dos processos centrais da citocinese. A totalidade do anel contrátil, o aparelho do fuso , os microtúbulos e o material periférico denso são chamados de "corpo de Fleming" ou "corpo intermediário".
- Apoptose . Durante a morte celular programada, a família de proteases ICE / ced-3 (uma das proteases do conversor de interleucina-1β) degradam a actina em dois fragmentos in vivo ; um dos fragmentos tem 15 kDa e o outro 31 kDa. Isso representa um dos mecanismos envolvidos na destruição da viabilidade celular que forma a base da apoptose. A protease calpaína também demonstrou estar envolvida neste tipo de destruição celular; assim como o uso de inibidores de calpaína demonstrou diminuir a proteólise da actina e a degradação do DNA (outro dos elementos característicos da apoptose). Por outro lado, o desencadeamento da apoptose induzida por estresse provoca a reorganização do citoesqueleto de actina (que também envolve sua polimerização), dando origem a estruturas chamadas fibras de estresse ; isso é ativado pela via da MAP quinase .

- Adesão e desenvolvimento celular . A adesão entre as células é uma característica dos organismos multicelulares que permite a especialização do tecido e, portanto, aumenta a complexidade celular. A adesão do epitélio celular envolve o citoesqueleto de actina em cada uma das células unidas, bem como as caderinas atuando como elementos extracelulares com a conexão entre os dois mediada por cateninas . Interferir na dinâmica da actina tem repercussões no desenvolvimento de um organismo, na verdade a actina é um elemento tão crucial que sistemas de genes redundantes estão disponíveis. Por exemplo, se o gene α-actinina ou fator de gelificação foi removido em Dictyostelium, os indivíduos não apresentam um fenótipo anômalo, possivelmente devido ao fato de que cada uma das proteínas pode desempenhar a função da outra. No entanto, o desenvolvimento de mutações duplas que não possuem os dois tipos de genes é afetado.
- Modulação da expressão gênica . O estado de polimerização da actina afeta o padrão de expressão do gene . Em 1997, foi descoberto que a despolimerização mediada pela citocalasina D nas células de Schwann causa um padrão específico de expressão para os genes envolvidos na mielinização desse tipo de célula nervosa . Foi demonstrado que a F-actina modifica o transcriptoma em alguns dos estágios da vida de organismos unicelulares, como o fungo Candida albicans . Além disso, proteínas semelhantes à actina desempenham um papel regulador durante a espermatogênese em camundongos e, em leveduras, acredita-se que as proteínas semelhantes à actina desempenhem um papel na regulação da expressão gênica . Na verdade, a actina é capaz de atuar como um iniciador da transcrição quando reage com um tipo de miosina nuclear que interage com RNA polimerases e outras enzimas envolvidas no processo de transcrição.
- Dinâmica de estereocílios . Algumas células desenvolvem protuberâncias filiformes finas em sua superfície que têm uma função mecanossensorial . Por exemplo, este tipo de organela está presente no Órgão de Corti , que está localizado no ouvido . A principal característica dessas estruturas é que seu comprimento pode ser modificado. A arquitetura molecular dos estereocílios inclui um núcleo paracristalino de actina em equilíbrio dinâmico com os monômeros presentes no citosol adjacente. As miosinas tipo VI e VIIa estão presentes em todo este núcleo, enquanto a miosina XVa está presente em suas extremidades em quantidades proporcionais ao comprimento dos estereocílios.
- Quiralidade intrínseca . Redes de actomiosina têm sido implicadas na geração de uma quiralidade intrínseca em células individuais. As células que crescem em superfícies quirais podem apresentar uma tendência direcional esquerda / direita dependente da actomiosina.
Patologia molecular
A maioria dos mamíferos possui seis genes de actina diferentes . Destes, dois codificam o citoesqueleto ( ACTB e ACTG1 ) enquanto os outros quatro estão envolvidos no músculo estriado esquelético ( ACTA1 ), tecido muscular liso ( ACTA2 ), músculos intestinais ( ACTG2 ) e músculo cardíaco ( ACTC1 ). A actina no citoesqueleto está envolvida nos mecanismos patogênicos de muitos agentes infecciosos , incluindo o HIV . A grande maioria das mutações que afetam a actina são mutações pontuais que têm um efeito dominante , com exceção de seis mutações envolvidas na miopatia nemalínica . Isso ocorre porque, em muitos casos, o mutante do monômero de actina atua como um "tampão", evitando o alongamento da F-actina.
Patologia associada a ACTA1
ACTA1 é o gene que codifica a isoforma αda actina, predominante nos músculos estriados esqueléticos humanos, embora também se expresse no músculo cardíaco e na glândula tireoide . Sua sequência de DNA consiste em sete exons que produzem cinco transcrições conhecidas. A maioria consiste em mutações pontuais que causam a substituição de aminoácidos . Em muitos casos, as mutações estão associadas a um fenótipo que determina a gravidade e o curso da doença.

As altera mutação da estrutura e função do músculo esquelético que produzem uma das três formas de miopatia : Tipo 3 miopatia nemalínica , miopatia congénita com um excesso de miofilamentos finas (cm) e miopatia congénita com o tipo de fibra desproporção (CMFTD). Também foram encontradas mutações que produzem miopatias centrais . Embora seus fenótipos sejam semelhantes, além da miopatia nemalínica típica, alguns especialistas distinguem outro tipo de miopatia denominado miopatia nemalínica actínica. No primeiro caso, aglomerados de actina formam-se em vez dos bastonetes típicos. É importante ressaltar que um paciente pode apresentar mais de um desses fenótipos em uma biópsia . Os sintomas mais comuns consistem em morfologia facial típica ( fácies miopática ), fraqueza muscular, atraso no desenvolvimento motor e dificuldades respiratórias. O curso da doença, sua gravidade e a idade em que aparece são variáveis e também são encontradas formas sobrepostas de miopatia. Um sintoma de miopatia nemalínica é que os "bastonetes de nemalina" aparecem em locais diferentes nas fibras musculares do tipo 1. Esses bastonetes são estruturas não patognomônicas que possuem uma composição semelhante aos discos Z encontrados no sarcômero .
A patogênese dessa miopatia é muito variada. Muitas mutações ocorrem na região de indentação da actina perto de seus locais de ligação de nucleotídeos , enquanto outras ocorrem no Domínio 2, ou nas áreas onde ocorre interação com proteínas associadas. Isso explica de alguma forma a grande variedade de aglomerados que se formam nesses casos, como Nemaline, Intranuclear Bodies ou Zebra Bodies. Mudanças no dobramento da actina ocorrem na miopatia nemalínica, assim como mudanças em sua agregação e também mudanças na expressão de outras proteínas associadas. Em algumas variantes onde os corpos intranucleares são encontrados, as mudanças na dobra mascaram o sinal de exportação da proteína do núcleo, de modo que o acúmulo da forma mutada da actina ocorre no núcleo da célula . Por outro lado, parece que mutações em ACTA1 que dão origem a um CFTDM têm um efeito maior na função sarcomérica do que em sua estrutura. Investigações recentes tentaram entender esse aparente paradoxo, que sugere não haver correlação clara entre o número de hastes e a fraqueza muscular. Parece que algumas mutações são capazes de induzir uma maior taxa de apoptose nas fibras musculares do tipo II.

No músculo liso
Existem duas isoformas que codificam para actinas no tecido muscular liso :
ACTG2 codifica a maior isoforma de actina, que tem nove exons , um dos quais, o que está localizado na extremidade 5 ', não é traduzido . É uma γ-actina expressa no músculo liso entérico. Não foram encontradas mutações nesse gene que correspondam a patologias, embora microarrays tenham mostrado que essa proteína é mais frequentemente expressa em casos que são resistentes à quimioterapia com cisplatina .
ACTA2 codifica uma α-actina localizada no músculo liso e também no músculo liso vascular. Foi observado que a mutação MYH11 pode ser responsável por pelo menos 14% dos aneurismas hereditários da aorta torácica, particularmente do Tipo 6. Isso ocorre porque a variante mutada produz uma montagem filamentar incorreta e uma capacidade reduzida de contração do músculo liso vascular. A degradação da mídia aórtica foi registrada nesses indivíduos, com áreas de desorganização e hiperplasia , bem como estenose dos vasa vasorum da aorta. O número de aflições nas quais o gene está implicado está aumentando. Tem sido relacionada à doença de Moyamoya e parece provável que certas mutações em heterozigose possam conferir uma predisposição a muitas patologias vasculares, como aneurisma da aorta torácica e doença isquêmica do coração . A α-actina encontrada na musculatura lisa também é um marcador interessante para avaliar a evolução da cirrose hepática.
No músculo cardíaco
O gene ACTC1 codifica a isoforma α-actina presente no músculo cardíaco. Ele foi sequenciado pela primeira vez por Hamada e colegas de trabalho em 1982, quando foi descoberto que é interrompido por cinco introns. Foi o primeiro dos seis genes em que foram encontrados alelos que estavam implicados em processos patológicos.

Um número de perturbações estruturais associados com mutações pontuais neste gene têm sido descrito que causa o mau funcionamento do coração, tal como Tipo 1R cardiomiopatia dilatada e de Tipo 11, cardiomiopatia hipertrófica . Certos defeitos do septo atrial foram descritos recentemente que também podem estar relacionados a essas mutações.
Dois casos de cardiomiopatia dilatada tem sido estudada envolvendo uma substituição de altamente conservada de amino ácidos pertencentes aos domínios de proteínas que se ligam e intersperse com os discos Z . Isso levou à teoria de que a dilatação é produzida por um defeito na transmissão da força contrátil nos miócitos .
As mutações em ACTC1 são responsáveis por pelo menos 5% das cardiomiopatias hipertróficas. A existência de uma série de mutações pontuais também foi encontrada:
- Mutação E101K: mudanças de carga líquida e formação de uma ligação eletrostática fraca no local de ligação da actomiosina.
- P166A: zona de interação entre monômeros de actina.
- A333P: zona de interação actina-miosina.
A patogênese parece envolver um mecanismo compensatório: as proteínas mutantes agem como toxinas com efeito dominante, diminuindo a capacidade do coração de se contrair, causando comportamento mecânico anormal de tal forma que a hipertrofia, que geralmente é retardada, é uma consequência da resposta normal do músculo cardíaco ao estresse .
Estudos recentes descobriram mutações ACTC1 que estão implicadas em dois outros processos patológicos: cardiomiopatia restritiva idiopática infantil e miocárdio não compactado do ventrículo esquerdo .
Em actinas citoplasmáticas
ACTB é um locus altamente complexo. Existemvários pseudogenes que são distribuídos por todo o genoma , e sua sequência contém seis exons que podem dar origem a até 21 transcrições diferentes por splicing alternativo , que são conhecidas como β-actinas. Consistente com esta complexidade, seus produtos também se encontram em vários locais e fazem parte de uma grande variedade de processos ( citoesqueleto ,complexo NuA4 histona -aciltransferase, núcleo celular ) e, além disso, estão associados a mecanismos de um grande número de processos patológicos ( carcinomas , distonia juvenil, mecanismos de infecção,malformações do sistema nervoso e invasão tumoral, entre outros). Uma nova forma de actina foi descoberta, kappa actina, que parece substituir a β-actina em processos relacionados a tumores .

Três processos patológicos foram descobertos até agora que são causados por uma alteração direta na sequência do gene:
- Hemangiopericitoma com t (7; 12) (p22; q13) -translocations é uma doença rara, em que uma mutação translocational provoca a fusão do ACTB gene sobre GLI1 no cromossoma 12 .
- A distonia de início juvenil é uma doença degenerativa rara que afeta o sistema nervoso central ; em particular, afeta áreas do neocórtex e tálamo , onde inclusões eosinofílicas semelhantes a bastonetes são formadas. Os indivíduos afetados representam um fenótipo com deformidades na linha mediana, hipoacusia sensorial e distonia. É causada por uma mutação pontual na qual o aminoácido triptofano substitui a arginina na posição 183. Isso altera a interação da actina com o sistema ADF / cofilina , que regula a dinâmica da formação do citoesqueleto das células nervosas .
- Também foi descoberta uma mutação de ponto dominante que causa disfunção dos granulócitos neutrófilos e infecções recorrentes . Parece que a mutação modifica o domínio responsável pela ligação entre a profilina e outras proteínas reguladoras. A afinidade da actina pela profilina é bastante reduzida neste alelo.
O locus ACTG1 codifica a proteína γ-actina citosólica que é responsável pela formação dos microfilamentos do citoesqueleto . Ele contém seis exons , dando origem a 22 mRNAs diferentes , que produzem quatro isoformas completas, cuja forma de expressão provavelmente depende do tipo de tecido em que se encontram. Também tem dois promotores de DNA diferentes . Notou-se que as sequências traduzidas desse locus e daquele da β-actina são muito semelhantes às previstas, sugerindo uma sequência ancestral comum que sofreu duplicação e conversão genética.
Do ponto de vista patológico, está associada a processos como amiloidose , retinite pigmentosa , mecanismos de infecção, doenças renais e diversos tipos de surdez congênita.
Descobriu-se que seis mutações pontuais autossômicas dominantes na sequência causam vários tipos de perda auditiva, particularmente perda auditiva neurossensorial ligada ao locus DFNA 20/26. Parece que afetam os estereocílios das células ciliadas presentes no órgão de Corti do ouvido interno . A β-actina é a proteína mais abundante encontrada no tecido humano, mas não é muito abundante nas células ciliadas, o que explica a localização da patologia. Por outro lado, parece que a maioria dessas mutações afeta as áreas envolvidas na ligação com outras proteínas, particularmente a actomiosina. Alguns experimentos sugeriram que o mecanismo patológico para esse tipo de perda auditiva está relacionado ao fato de a F-actina nas mutações ser mais sensível à cofilina do que o normal.
No entanto, embora não haja registro de nenhum caso, sabe-se que a γ-actina também se expressa nos músculos esqueléticos e, embora esteja presente em pequenas quantidades, organismos modelo demonstraram que sua ausência pode dar origem a miopatias.
Outros mecanismos patológicos
Alguns agentes infecciosos usam actina, especialmente actina citoplasmática, em seu ciclo de vida . Duas formas básicas estão presentes nas bactérias :
- Listeria monocytogenes , algumas espécies de Rickettsia , Shigella flexneri e outros germes intracelulares escapam dosvacúolos fagocíticos revestindo-se com uma cápsula de filamentos de actina. L. monocytogenes e S. flexneri geram uma cauda na forma de uma "cauda de cometa" que lhes dá mobilidade. Cada espécie exibe pequenas diferenças no mecanismo de polimerização molecular de suas "caudas de cometa". Diferentes velocidades de deslocamento foram observadas, por exemplo, com Listeria e Shigella consideradas as mais rápidas. Muitos experimentos demonstraram esse mecanismo in vitro . Isso indica que a bactéria não está usando um motor de proteína semelhante à miosina e parece que sua propulsão é adquirida a partir da pressão exercida pela polimerização que ocorre próximo à parede celular do microrganismo. A bactéria foi anteriormente envolvida por ABPs do hospedeiro e, no mínimo, a cobertura contém complexo Arp2 / 3 , proteínas Ena / VASP , cofilina, uma proteína tampão e promotores de nucleação, como ocomplexo vinculina . Por meio desses movimentos, eles formam protuberâncias que atingem as células vizinhas, infectando-as também para que o sistema imunológico só possa combater a infecção por meio da imunidade celular. O movimento pode ser causado pela modificação da curva e desramificação dos filamentos. Outras espécies, como Mycobacterium marinum e Burkholderia pseudomallei , também são capazes de polimerização localizada da actina celular para auxiliar seu movimento por meio de um mecanismo que está centrado no complexo Arp2 / 3. Além disso, o vírus da vacina Vaccinia também usa elementos do citoesqueleto de actina para sua disseminação.
- Pseudomonas aeruginosa é capaz de formar um biofilme protetorpara escapardas defesas deum organismo hospedeiro , especialmente glóbulos brancos e antibióticos . O biofilme é construído usandofilamentos de DNA e actina do organismo hospedeiro.
Além do exemplo citado anteriormente, a polimerização da actina é estimulada nas etapas iniciais da internalização de alguns vírus, notadamente HIV , por exemplo, inativando o complexo de cofilina.
O papel que a actina desempenha no processo de invasão das células cancerosas ainda não foi determinado.
Evolução
O citoesqueleto eucariótico de organismos entre todos os grupos taxonômicos tem componentes semelhantes à actina e tubulina. Por exemplo, a proteína que é codificada pelo gene ACTG2 em humanos é completamente equivalente aos homólogos presentes em ratos e camundongos, embora em um nível de nucleotídeo a similaridade diminua para 92%. No entanto, existem grandes diferenças com os equivalentes em procariotos ( FtsZ e MreB ), onde a semelhança entre as sequências de nucleotídeos está entre 40-50% entre as diferentes bactérias e espécies de archaea . Alguns autores sugerem que a proteína ancestral que deu origem ao modelo da actina eucariótica se assemelha às proteínas presentes nos citoesqueletos bacterianos modernos.

Alguns autores apontam que o comportamento da actina, tubulina e histona , uma proteína envolvida na estabilização e regulação do DNA, são semelhantes em sua capacidade de ligar nucleotídeos e em sua capacidade de aproveitar o movimento browniano . Também foi sugerido que todos eles têm um ancestral comum. Portanto, os processos evolutivos resultaram na diversificação das proteínas ancestrais nas variedades atuais, conservando, entre outras, as actinas como moléculas eficientes que eram capazes de atacar processos biológicos ancestrais essenciais, como a endocitose .
Equivalentes em bactérias
O citoesqueleto bacteriano pode não ser tão complexo quanto o encontrado nos eucariotos ; no entanto, ele contém proteínas que são altamente semelhantes aos monômeros e polímeros de actina. A proteína bacteriana MreB polimeriza em finos filamentos não helicoidais e, ocasionalmente, em estruturas helicoidais semelhantes à F-actina. Além disso, sua estrutura cristalina é muito semelhante à da G-actina (em termos de sua conformação tridimensional), existem até semelhanças entre os protofilamentos MreB e F-actina. O citoesqueleto bacteriano também contém as proteínas FtsZ , que são semelhantes à tubulina .
As bactérias, portanto, possuem um citoesqueleto com elementos homólogos à actina (por exemplo, MreB, AlfA, ParM , FtsA e MamK), embora a sequência de aminoácidos dessas proteínas seja diferente daquela presente nas células animais. No entanto, essas proteínas têm um alto grau de semelhança estrutural com a actina eucariótica. Os microfilamentos altamente dinâmicos formados pela agregação de MreB e ParM são essenciais para a viabilidade celular e estão envolvidos na morfogênese celular, segregação cromossômica e polaridade celular. ParM é um homólogo de actina que é codificado em um plasmídeo e está envolvido na regulação do DNA do plasmídeo. ParMs de diferentes plasmídeos bacterianos podem formar estruturas helicoidais surpreendentemente diversas compreendendo duas ou quatro fitas para manter a herança fiel do plasmídeo.
Formulários
A actina é usada em laboratórios científicos e tecnológicos como uma pista para motores moleculares como a miosina (no tecido muscular ou fora dele) e como um componente necessário para o funcionamento celular. Também pode ser utilizado como ferramenta diagnóstica, pois várias de suas variantes anômalas estão relacionadas ao aparecimento de patologias específicas.
- Nanotecnologia . Os sistemas actina-miosina atuam como motores moleculares que permitem o transporte de vesículas e organelas por todo o citoplasma. É possível que a actina possa ser aplicada à nanotecnologia, visto que sua capacidade dinâmica foi aproveitada em uma série de experimentos, incluindo aqueles realizados em sistemas acelulares. A ideia subjacente é usar os microfilamentos como trilhas para guiar os motores moleculares que podem transportar uma determinada carga. Ou seja, a actina pode ser usada para definir um circuito ao longo do qual uma carga pode ser transportada de uma maneira mais ou menos controlada e direcionada. Em termos de aplicações gerais, poderia ser utilizado para o transporte direcionado de moléculas para depósito em determinados locais, o que permitiria a montagem controlada de nanoestruturas. Esses atributos podem ser aplicados a processos de laboratório, como em lab-on-a-chip , em mecânica de nanocomponentes e em nanotransformadores que convertem energia mecânica em energia elétrica.

- A actina é usada como um controle interno em western blots para verificar se quantidades iguais de proteína foram carregadas em cada faixa do gel. No exemplo de transferência mostrado no lado esquerdo, 75 µg de proteína total foram carregados em cada poço. O blot reagiu com anticorpo anti-β-actina (para outros detalhes do blot, consulte a referência)
O uso da actina como controle interno é baseado no pressuposto de que sua expressão é praticamente constante e independente das condições experimentais. Ao comparar a expressão do gene de interesse com a da actina, é possível obter uma quantidade relativa que pode ser comparada entre diferentes experimentos, sempre que a expressão desta for constante. Vale ressaltar que nem sempre a actina apresenta a estabilidade desejada na expressão gênica .
- Saúde. Alguns alelos da actina causam doenças; por este motivo foram desenvolvidas técnicas para a sua detecção. Além disso, a actina pode ser utilizada como marcador indireto em patologia cirúrgica: é possível utilizar variações no padrão de sua distribuição nos tecidos como marcador de invasão em neoplasias , vasculites e outras condições. Além disso, devido à estreita associação da actina com o aparelho de contração muscular, seus níveis no músculo esquelético diminuem quando esses tecidos atrofiam , ela pode, portanto, ser usada como um marcador desse processo fisiológico.
- Tecnologia de alimentos . É possível determinar a qualidade de certos alimentos processados, como salsichas , por meio da quantificação da quantidade de actina presente na carne constituinte. Tradicionalmente, tem-se utilizado um método baseado na detecção de 3-metilhistidina em amostras hidrolisadas desses produtos, uma vez que esse composto está presente na cadeia pesada da actina e da F-miosina (ambas são componentes principais do músculo). A geração desse composto na carne deriva da metilação dos resíduos de histidina presentes em ambas as proteínas.
Genes
Os genes humanos que codificam proteínas actinas incluem:
Veja também
- Remodelação de actina - efeito na estrutura e forma celular
- Proteína de ligação à actina
- Matéria ativa
- Arp2 / 3
- Filopodia
- FtsZ
- Filamento intermediário
- Lamelipódio
- Proteína motora - converte energia química em trabalho mecânico
- MreB - um dos homólogos da actina em bactérias
- Neurônio
- Falotoxina
Referências
links externos
- Técnicas de coloração com actina (coloração de células vivas e fixas)
- Classe de motivo de recurso de Motivo Linear Eucariótico LIG_Actin_RPEL_3
- Classe de motivo de recurso de Motivo Linear Eucariótico LIG_Actin_WH2_1
- Classe de motivo de recurso de Motivo Linear Eucariótico LIG_Actin_WH2_2
- Estruturas macromoleculares 3D de filamentos de actina do Banco de Dados EM (EMDB)