
Seymouria -Seymouria
|
Seymouria |
|
|---|---|

|
|
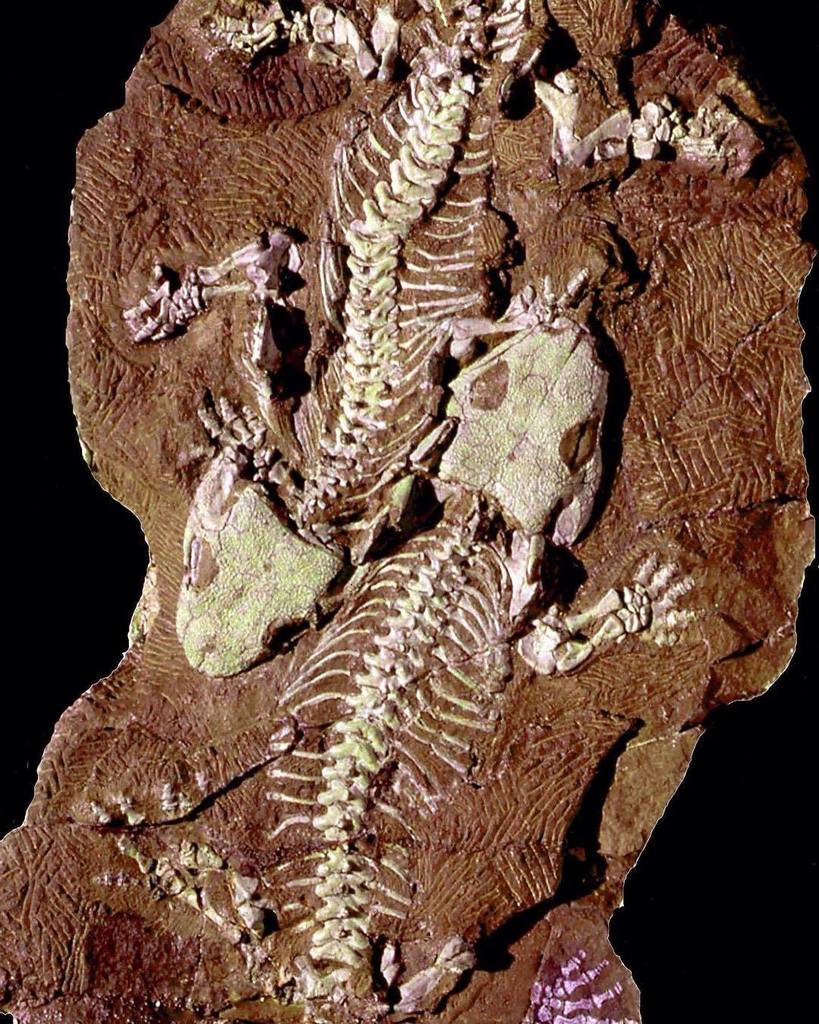
| Um fóssil de Seymouria baylorensis exibido no Museu Nacional de História Natural | |
|
Classificação científica |
|
| Reino: | Animalia |
| Filo: | Chordata |
| Pedido: | † Seymouriamorpha |
| Família: | † Seymouriidae |
| Gênero: |
† Seymouria Broili, 1904 |
| Espécies | |
|
|
| Sinônimos | |
|
|
Seymouria é um gênero extinto de Seymouriamorph do início do Permiano da América do Norte e Europa . Embora fossem anfíbios (no sentido biológico), Seymouria estava bem adaptado à vida na terra, com muitascaracterísticas reptilianas - tantas, na verdade, que Seymouria foi inicialmente considerado um réptil primitivo. É conhecido principalmente por duas espécies, Seymouria baylorensis e Seymouria sanjuanensis . A espécie-tipo, S. baylorensis , é mais robusta e especializada, embora seus fósseis só tenham sido encontrados no Texas . Por outro lado, Seymouria sanjuanensis é mais abundante e difundida. Esta espécie menor é conhecida por vários fósseis bem preservados, incluindo um bloco de seis esqueletos encontrados na Formação Cutler do Novo México e um par de esqueletos totalmente crescidos da Formação Tambach da Alemanha , que foram fossilizados um ao lado do outro.
Durante a primeira metade do século 20, Seymouria foi considerado um dos répteis mais antigos e "primitivos" conhecidos. Os paleontólogos observaram como a forma geral do corpo se assemelhava à dos primeiros répteis, como os captorinídeos , e que certas adaptações dos membros, quadril e crânio também eram semelhantes às dos primeiros répteis, em vez de qualquer espécie de anfíbios modernos ou extintos conhecidos na época . Os membros e espinha dorsal fortemente construídos também apoiavam a ideia de que Seymouria era principalmente terrestre, passando muito pouco tempo na água. No entanto, na década de 1950, girinos fossilizados foram descobertos em Discosauriscus , que era um parente próximo de Seymouria no grupo Seymouriamorpha . Isso mostra que seymouriamorphs (incluindo Seymouria ) tinham um estágio larval que vivia na água, tornando Seymouria não um verdadeiro réptil, mas sim um anfíbio que estava intimamente relacionado aos répteis. Embora não seja mais considerado o réptil mais primitivo, Seymouria ainda é um fóssil transicional importante que documenta a aquisição de características esqueléticas semelhantes a répteis antes da evolução do ovo amniótico, que caracteriza os amniotas (répteis, mamíferos e pássaros ).
História

Os fósseis de Seymouria foram encontrados pela primeira vez perto da cidade de Seymour , no condado de Baylor , Texas (daí o nome da espécie-tipo, Seymouria baylorensis , referindo-se tanto à cidade quanto ao condado). Os primeiros fósseis a serem descobertos foram um agrupamento de indivíduos coletados por CH Sternberg em 1882. No entanto, esses fósseis não seriam devidamente preparados e identificados como Seymouria até 1930.
Vários paleontólogos de todo o mundo recuperaram seus próprios fósseis de Seymouria baylorensis no final do século 19 e no início do século 20. Os primeiros fósseis explicitamente nomeados como Seymouria eram um par de crânios incompletos, um dos quais foi preservado com alguns elementos peitorais e vertebrais. Esses fósseis foram descritos pelo paleontólogo alemão Ferdinand Broili em 1904 e agora estão armazenados em Munique . O paleontólogo americano SW Williston mais tarde descreveu um esqueleto quase completo em 1911, e notou que " Desmospondylus anomalus ", um táxon que ele havia batizado recentemente a partir de membros fragmentados e vértebras, provavelmente representava indivíduos juvenis ou mesmo embrionários de Seymouria .
Da mesma forma, o paleontólogo inglês DMS Watson observou em 1918 que Conodectes , um gênero duvidoso nomeado pelo famoso Edward Drinker Cope em 1896, era provavelmente sinônimo de Seymouria . Robert Broom (1922) argumentou que o gênero deveria ser referido como Conodectes desde que o nome foi publicado primeiro, mas Alfred Romer (1928) objetou, observando que o nome Seymouria era muito popular na comunidade científica para ser substituído. Durante esse tempo, Seymouria era geralmente visto como um réptil muito antigo, parte de um grau evolutivo conhecido como "cotilossauros", que também incluía muitos outros répteis do Permiano de corpo robusto ou tetrápodes semelhantes a répteis. No entanto, muitos paleontólogos não tinham certeza sobre sua lealdade aos répteis, notando muitas semelhanças com os embolômeros , que eram inquestionavelmente anfíbios "labirintodontes". Esta combinação de características de répteis (ou seja, outros "cotilossauros") e anfíbios (ou seja, embolômeros) foi a evidência de que Seymouria foi fundamental para a transição evolutiva entre os dois grupos. Apesar de tudo, não se sabia o suficiente sobre sua biologia para concluir de qual grupo ele realmente fazia parte. Broom (1922) e o paleontólogo russo Peter Sushkin (1925) apoiaram uma colocação entre os anfíbios, mas a maioria dos estudos nessa época provisoriamente considerou-o um réptil extremamente "primitivo"; estes incluíram uma redescrição abrangente do material referido à espécie, publicado por Theodore E. White em 1939.

No entanto, começaram a surgir evidências indiretas de que Seymouria não era biologicamente reptiliano. Na década de 1940, vários gêneros foram ligados a Seymouria como parte do grupo Seymouriamorpha . Alguns seymouriamorphs, como o Kotlassia , tinham evidências de hábitos aquáticos, e até mesmo o próprio Seymouria às vezes era acusado de possuir linhas laterais, estruturas sensoriais utilizáveis apenas debaixo d'água. Watson (1942) e Romer (1947) inverteram cada um sua posição sobre a classificação de Seymouria , colocando-a entre os anfíbios e não entre os répteis. Talvez a evidência mais contundente tenha surgido em 1952, quando o paleontólogo tcheco Zdeněk Špinar relatou guelras preservadas em fósseis juvenis do seymouriamorph Discosauriscus . Isso provou inequivocamente que os seymouriamorphs tinham um estágio larval e, portanto, eram anfíbios, biologicamente falando. No entanto, a grande quantidade de semelhanças entre Seymouria e répteis apoiava a ideia de que os seymouriamorphs eram bastante próximos à ancestralidade dos amniotas .

Em 1966, Peter Paul Vaughn descreveu os restos mortais de Seymouria do xisto de Organ Rock de Utah . Esses restos, que eram uma variedade de crânios, representavam uma nova espécie, Seymouria sanjuanensis . Descobriu-se que os fósseis desta espécie são mais abundantes e difundidos do que os de Seymouria baylorensis . Várias outras espécies foram posteriormente nomeadas por Paul E. Olson , embora sua validade tenha sido mais questionável do que a de S. sanjuanensis . Por exemplo, Seymouria agilis (Olson, 1980), conhecido a partir de um esqueleto quase completo da Formação Chickasha de Oklahoma , foi reatribuído por Michel Laurin e Robert R. Reisz ao macróleter parareptil em 2001. Seymouria grandis , descrita um ano antes de um A caixa craniana encontrada no Texas não foi referenciada a nenhum outro tetrápode, mas permanece pouco conhecida. Langston (1963) relatou um fêmur indistinguível daquele de S. baylorensis em sedimentos Permianos na Ilha do Príncipe Eduardo, na costa leste do Canadá . Restos de esqueletos semelhantes a Seymouria também são conhecidos na pedreira de Richards Spur em Oklahoma, conforme descrito pela primeira vez por Sullivan & Reisz (1999).
Um bloco de sedimento contendo seis esqueletos de S. sanjuanensis foi encontrado na Formação Cutler do Novo México , conforme descrito por Berman, Reisz e Eberth (1987). Em 1993, Berman & Martens relataram os primeiros vestígios de Seymouria fora da América do Norte , quando descreveram fósseis de S. sanjuanensis da Formação Tambach da Alemanha . A Formação Tambach também produziu fósseis de S. sanjuanensi de qualidade semelhante aos da Formação Cutler. Por exemplo, em 2000, Berman e seus colegas descreveram os "Amantes Tambach", dois esqueletos completos e totalmente articulados de S. sanjuanensis fossilizados lado a lado (embora não seja possível determinar se eles eram um casal morto durante o acasalamento). A Formação Tambach também produziu os fósseis conhecidos mais novos em desenvolvimento de Seymouria , auxiliando nas comparações com Discosauriscus , que é conhecido principalmente em juvenis.
Descrição
Os indivíduos de Seymouria eram animais de constituição robusta, com cabeça grande, pescoço curto, membros atarracados e pés largos. Eles eram bastante pequenos, apenas cerca de 2 pés (60 cm) de comprimento. O crânio era quadradão e aproximadamente triangular quando visto de cima, mas era mais baixo e mais longo do que a maioria dos outros seymouriamorphs. As vértebras tinham arcos neurais amplos e inchados (a parte acima da medula espinhal). Como um todo, a forma do corpo era semelhante à dos répteis contemporâneos e dos tetrápodes semelhantes aos répteis, como os captorinídeos , diadectomorfos e pararepteis . Coletivamente, esses tipos de animais foram chamados de "cotilossauros" no passado, embora não formem um clado (um agrupamento natural baseado em relações).
Crânio

O crânio era composto de muitos ossos menores em forma de placa. A configuração dos ossos do crânio presentes em Seymouria era muito semelhante à dos tetrápodes e parentes dos tetrápodes muito mais antigos. Por exemplo, ele retém um osso intertemporal , que é a condição plesiomórfica ("primitiva") presente em animais como Ventastega e embolômeros . Os ossos do crânio eram fortemente texturizados, como era típico dos antigos anfíbios e répteis captorinídeos . Além disso, a parte posterior do crânio apresentava uma grande incisão que se estendia ao longo de sua lateral. Essa incisão foi denominada entalhe ótico , e uma incisão semelhante na mesma área geral é comum à maioria dos anfíbios paleozóicos ("labirintodontes", como às vezes são chamados), mas desconhecida nos amniotas. A borda inferior da incisura ótica era formada pelo osso esquamosal , enquanto a borda superior era formada por flanges voltados para baixo dos ossos supratemporal e tabular (conhecidos como flanges óticos). A tabular também tem uma segunda flange voltada para baixo, visível da parte de trás do crânio; esta flange (conhecida como flange occipital) conectava-se à caixa craniana e obscurecia parcialmente o espaço entre a caixa craniana e a lateral do crânio. O desenvolvimento das flanges óticas e occipitais é maior em Seymouria (particularmente S. baylorensis ) do que em qualquer outro seymouriamorph.
O aparato sensorial do crânio também merece menção por uma série de características únicas. As órbitas (órbitas) ficavam no meio do comprimento do crânio, embora estivessem um pouco mais próximas do focinho nos juvenis. Eles eram mais romboidais do que as órbitas circulares de outros seymouriamorphs, com uma borda frontal aguda. Vários autores notaram que alguns espécimes de Seymouria possuíam sulcos indistintos, presentes nos ossos ao redor das órbitas e na frente da incisura ótica. Esses sulcos eram provavelmente remanescentes de um sistema de linha lateral , uma teia de órgãos sensores de pressão úteis para animais aquáticos, incluindo o presumível estágio larval de Seymouria . Muitos espécimes não retêm nenhum vestígio de suas linhas laterais, nem mesmo os juvenis. Aproximadamente no meio dos ossos parietais havia um pequeno orifício conhecido como forame pineal, que pode ter contido um órgão sensorial conhecido como olho parietal . O forame pineal é menor em Seymouria do que em outros seymouriamorphs.
O estribo , um osso em forma de haste que fica entre a caixa craniana e a parede do crânio, era cônico. Ele conectava a caixa craniana à borda superior do entalhe ótico e provavelmente servia como um canal de vibrações recebidas por um tímpano (tímpano) que presumivelmente estava dentro do entalhe ótico. Dessa forma, ele pode transmitir o som do mundo externo para o cérebro. A configuração do estribo é intermediária entre os tetrápodes não amniotas e os amniotas. Por um lado, sua conexão com a incisura ótica é incomum, uma vez que répteis verdadeiros e outros amniotas perderam uma incisura ótica, forçando o tímpano e o estribo a se deslocarem para baixo em direção ao osso quadrático da articulação da mandíbula. Por outro lado, a estrutura fina e sensível do estribo de Seymouria é uma especialização sobre a maioria dos tetrápodes não amniotas, que possuem estribo espesso, inútil para a audição, mas útil para reforçar o crânio. O ouvido interno de Seymouria baylorensis retém um recesso coclear localizado atrás (em vez de abaixo) do vestíbulo , e seu canal semicircular anterior era provavelmente circundado por um supraoccipital cartilaginoso (em vez de ósseo) . Essas características são mais primitivas do que as dos verdadeiros répteis e sinapsídeos.
O palato (céu da boca) tinha algumas semelhanças com os tetrápodes amniotas e não amniotas. Por um lado, ele reteve algumas presas isoladas com dobras internas de esmalte labirínticas, como é característico dos anfíbios "labirintodontes". Por outro lado, os ossos do vômer na parte frontal da boca eram bastante estreitos e as coanas adjacentes (orifícios que conduziam da cavidade nasal à boca) eram grandes e próximos, como nos amniotas. O palato é geralmente de osso sólido, com apenas vacuidades interpteróides vestigiais (um par de orifícios ao longo da linha média) separados por um processo cultriforme longo e fino (o ramo anterior da base da caixa craniana). Além das presas isoladas, o palato também é coberto por pequenos dentículos que se irradiam da parte posterior dos ossos pterigóides . Seymouria tem algumas características do palato semelhantes a amniotas, como a presença de um ramo do pterigóide semelhante a uma ponta (formalmente conhecido como flange transversa), bem como um osso epipterigóide que é separado do pterigóide. No entanto, essas características foram observadas em vários tetrápodes não amniotas, portanto, não significam seu status de amniota.
A mandíbula inferior manteve algumas características plesiomórficas. Por exemplo, a borda interna da mandíbula possui três ossos coronóides. A mandíbula também reteve pelo menos um grande orifício ao longo de sua borda interna, conhecido como fenestra meckeliana, embora essa característica só tenha sido confirmada durante uma nova investigação em 2005 de um dos espécimes da Formação Cutler. Nenhuma dessas características é comum em amniotas. A caixa craniana tinha um mosaico de características em comum com vários tetrapodomorfos. O sistema de sulcos e aberturas nervosas na lateral da caixa craniana eram incomumente semelhantes aos dos peixes Megalichthys , e a base cartilaginosa é outra característica plesiomórfica. No entanto, as artérias carótidas internas perfuram a caixa craniana perto da parte posterior do complexo ósseo, uma característica derivada semelhante aos amniotas.
Esqueleto pós-craniano

A coluna vertebral é bastante curta, com um total de 24 vértebras na frente do quadril. As vértebras são gastrocêntricas, o que significa que cada vértebra tem um componente maior, em forma de carretel, conhecido como pleurocentrum, e um componente menor em forma de cunha (ou crescente na frente) conhecido como intercentro. Os arcos neurais, que ficam acima do pleurocentro, são dilatados em estruturas largas com zigapófises semelhantes a uma mesa (placas articulares) cerca de três vezes mais largas que o próprio pleurocentro. Algumas vértebras têm espinhas neurais parcialmente subdivididas ao meio, enquanto outras são ovais em seção transversal. As costelas das vértebras dorsais se estendem horizontalmente e se fixam às vértebras em dois lugares: o intercentro e o lado do arco neural. O pescoço está praticamente ausente, apenas algumas vértebras de comprimento. A primeira vértebra cervical, o atlas , tinha um pequeno intercentro, bem como um pleurocentro reduzido, que só estava presente em indivíduos maduros. Embora o atlantal pleurocentrum (quando presente) foi em cunha entre o intercentrum do atlas e intercentrum do sucedendo eixo , o desenvolvimento ósseo baixo vértebra (como em amniotes) nesta área dos contrastes pescoço com os característicos atlas eixo complexo de amniotes. Além disso, estudos posteriores descobriram que o atlas intercentrum foi dividido em uma parte esquerda e direita, mais parecida com a dos tetrápodes do tipo anfíbio. Ao contrário de quase todos os outros tetrápodes paleozóicos (amniotas ou não), Seymouria carece completamente de quaisquer vestígios ósseos de escamas ou escamas, nem mesmo as escamas circulares da barriga de outros seymouriamorphs.
A cintura peitoral (ombro) tem várias características semelhantes a répteis. Por exemplo, a escápula e o coracóide (placas ósseas que ficam acima e abaixo da cavidade do ombro, respectivamente) são ossos separados, em vez de uma única omoplata grande. Da mesma forma, a interclavícula era plana e em forma de cogumelo, com uma "haste" longa e delgada. O úmero (osso do antebraço) tinha a forma de um L quadradão ligeiramente torcido, com grandes áreas para inserção do músculo. Esta forma, que foi descrita como "tetraédrica", é plesiomórfica para os tetrápodes. Por outro lado, o úmero também tem uma adaptação semelhante à de um réptil: um orifício conhecido como forame entepicondilar . O raio era aproximadamente em forma de ampulheta. A ulna é semelhante, mas mais longa devido à presença de um processo olécrano pronunciado , como é comum em tetrápodes terrestres, mas raro em anfíbios ou aquáticos. O carpo (pulso) tem dez ossos e a mão tem cinco dedos fortes. Os ossos do carpo estão totalmente desenvolvidos e em contato íntimo entre si, outra indicação de terrestrialidade. As falanges (ossos dos dedos) diminuem de tamanho em direção à ponta dos dedos, onde cada uma termina em um segmento minúsculo e arredondado, sem garra. A fórmula falangeal (número de falanges por dedo, do polegar ao mínimo) é 2-3-4-4-3.
Duas vértebras sacrais (quadril) estavam presentes, embora apenas a primeira possuísse uma costela grande e robusta que contatava o ílio (lâmina superior do quadril). Alguns estudos argumentaram que havia apenas uma vértebra sacral, com a suposta segunda sacral sendo na verdade a primeira caudal por ter uma costela mais curta e curva do que a primeira sacral. Cada ílio é baixo e em forma de lágrima quando visto de lado, enquanto a parte inferior do quadril como um todo é formada por uma única placa puboisquiática robusta, que é retangular quando vista de baixo. Ambas as órbitas do quadril e dos ombros foram direcionadas 45 graus abaixo da horizontal. O fêmur é tão robusto quanto o úmero, e a tíbia e a fíbula são ossos robustos em forma de ampulheta, semelhantes ao rádio e à ulna. O tarso (tornozelo) incorpora 11 ossos, intermediários entre os tetrápodes anteriores (que têm 12) e os amniotas (que têm 8 ou menos). Os pés de cinco dedos são bastante semelhantes às mãos, com fórmula falangeal 2-3-4-5-3.
Havia apenas cerca de 20 vértebras caudais (cauda) no máximo. Depois da base da cauda, os caudais começam a adquirir espinhos ósseos ao longo de sua parte inferior, conhecidos como divisas . Estes começam a aparecer nas proximidades do terceiro ao sexto caudal, dependendo do espécime. As costelas estão presentes apenas nas primeiras cinco ou seis caudais; eles são longos na base da cauda, mas diminuem logo depois e geralmente desaparecem em torno da mesma área em que as divisas aparecem.
Diferenças entre espécies
Seymouria baylorensis e Seymouria sanjuanensis podem ser distinguidas umas das outras com base em várias diferenças na forma e nas conexões entre os diferentes ossos do crânio. Por exemplo, a flange de osso voltada para baixo acima da incisura ótica (às vezes chamada de "chifre tabular" ou "processo ótico") é muito mais desenvolvida em S. baylorensis do que em S. sanjuanensis ; adquire uma forma triangular (quando vista de lado) à medida que se estende para baixo mais extensivamente em direção à parte posterior do crânio. Em S. sanjuanensis , o osso pós - frontal entra em contato com o osso parietal por meio de uma sutura obtusa em cunha, enquanto a conexão entre os dois ossos é completamente reta em S. baylorensis .

Alguns autores argumentaram que os pós- parietais de S. baylorensis eram menores do que os de S. sanjuanensis , mas alguns espécimes de S. sanjuanensis (por exemplo, os "amantes de Tambach") também tinham pós-parietais pequenos. Além disso, os "amantes de Tambach" têm um osso quadratojugal que é mais semelhante ao de S. baylorensis do que de S. sanjuanensis . A combinação de características de ambas as espécies nesses espécimes pode indicar que as duas espécies são parte de uma linhagem contínua, ao invés de dois caminhos evolutivos fortemente diferenciados. Da mesma forma, algumas diferenças relacionadas às proporções da parte posterior do crânio podem ser consideradas um artefato do fato de que a maioria dos espécimes de S. sanjuanensis não estavam totalmente crescidos antes da descoberta dos "amantes de Tambach", que eram membros adultos de as espécies.
No entanto, várias características ainda são claramente diferenciadas entre as duas espécies. O osso lacrimal , na frente dos olhos, ocupa apenas a borda frontal da órbita em S. baylorensis . No entanto, os espécimes de S. sanjuanensis têm um ramo lacrimal que se estende por uma pequena distância sob a órbita. Em S. sanjuanensis , grande parte da borda posterior da órbita é formada pelo osso pós-orbital em forma de chevron , que é mais retangular em S. baylorensis . A forma lacrimal e pós-orbitária de S. sanjuanensis corresponde intimamente à condição em outros seymouriamorphs, enquanto a condição em S. baylorensis é mais exclusiva e derivada.
O osso maxilar com dentes, que forma a lateral do focinho, também é distintamente único em S. baylorensis . Em S. sanjuanensis , a maxila era baixa, com muitos dentes afiados e bem espaçados estendendo-se ao longo de seu comprimento. Esta condição é semelhante a outros seymouriamorphs. No entanto, S. baylorensis tem um focinho mais alto e seus dentes são geralmente muito maiores, menos numerosos e menos homogêneos em tamanho. O palato é geralmente semelhante entre as duas espécies, embora os ectopterigóides sejam mais triangulares em S. baylorensis e retangulares em S. sanjuanensis .
Paleobiologia
Estilo de vida
Romer (1928) foi um dos primeiros autores a discutir as implicações biológicas do esqueleto de Seymouria . Ele argumentou que os membros robustos e o corpo largo sustentavam a ideia de que se tratava de um animal forte e terrestre com um andar amplo. No entanto, ele também observou que os rastros do Permiano geralmente apóiam a ideia de que os tetrápodes terrestres desse período não eram dragões de barriga, mas sim fortes o suficiente para manter seus corpos fora do chão. Assim como outros paleontólogos da época, Romer presumiu que Seymouria tinha um modo de reprodução reptiliano (ou amniota), com ovos postos em terra seca e protegidos dos elementos por uma membrana de âmnio .
White (1939) elaborou sobre as implicações biológicas. Ele observou que a presença de um entalhe ótico reduz a força da mandíbula, diminuindo a quantidade de área de superfície que os músculos da mandíbula podem se fixar dentro do crânio. Além disso, o crânio teria sido mais frágil devido à presença de uma incisão tão grande. Como um todo, ele achou improvável que Seymouria fosse capaz de atacar presas grandes e ativas. No entanto, os locais de inserção muscular no palato eram mais desenvolvidos do que os dos anfíbios contemporâneos. White extrapolou que Seymouria era um generalista carnívoro e onívoro, alimentando-se de invertebrados, pequenos peixes e talvez até mesmo algum material vegetal. Pode até ter sido canibal de acordo com seus cálculos.
White também chamou a atenção para o inchaço incomum das vértebras, o que teria facilitado o movimento lateral (lado a lado), mas proibia qualquer torção (torção) da coluna vertebral. Isso teria sido benéfico, já que Seymouria tinha membros baixos e um corpo largo e pesado que, de outra forma, seria vulnerável à torção quando estivesse andando. Isso também pode explicar a presença desse traço em captorinídeos, diadectomorfos e outros "cotilossauros". Talvez o inchaço das vértebras fosse uma estratégia provisória para evitar a torção, que mais tarde seria suplantada por fortes músculos do quadril nos répteis posteriores. Os músculos do quadril pouco desenvolvidos de Seymouria estão de acordo com esta hipótese. No entanto, essas vértebras eram ineficientes na defesa contra a torção em qualquer velocidade mais rápida do que uma caminhada rápida, então Seymouria provavelmente não era um animal de movimento rápido.
Embora White considerasse Seymoria bastante competente em terra, ele também discutiu alguns outros estilos de vida. Ele supôs que Seymouria também fosse um bom nadador, pois estimou (erroneamente) que o animal tinha uma cauda profunda e poderosa semelhante à dos crocodilianos modernos . No entanto, ele também observou que seria vulnerável a predadores semiaquáticos ou aquáticos, e que os fósseis de Seymouria eram mais comuns em depósitos terrestres como resultado de suas preferências de habitat. Berman et al . (2000) apoiou esta hipótese, já que a Formação Tambach preservou fósseis de Seymouria enquanto também carecia completamente de animais aquáticos. Eles também apontaram os ossos do pulso e do tornozelo bem desenvolvidos dos "amantes de Tambach" como suporte das afinidades terrestres. Apesar da forte musculatura dos membros anteriores, Romer (1928) e White (1939) encontraram poucas evidências de adaptações de escavação em Seymouria .
Dimorfismo sexual
Alguns autores argumentaram a favor do dimorfismo sexual existente em Seymouria , mas outros não estão convencidos por esta hipótese. White (1939) argumentou que alguns espécimes de Seymouria baylorensis tinham chevrons (espinhos ósseos na parte inferior das vértebras da cauda) que apareceram pela primeira vez na terceira vértebra da cauda, enquanto outros espécimes os tiveram pela primeira vez na sexta. Ele postulou que o aparecimento posterior das divisas em alguns espécimes era um indicativo de que eram machos que precisavam de mais espaço para armazenar sua genitália interna. Esse tipo de diferenciação sexual foi relatado tanto em tartarugas quanto em crocodilianos. Com base nisso, ele também apoiou a ideia de que as fêmeas Seymouria davam à luz ovos com gema grossa em terra, assim como as tartarugas e os crocodilianos. Vaughn (1966) mais tarde encontrou uma correlação entre a aquisição em chevron e certas proporções do crânio em Seymouria sanjuanensis , e propôs que eles também eram exemplos de dimorfismo sexual.
No entanto, Berman, Reisz, & Elberth (1987) criticaram as metodologias de White (1939) e Vaughn (1966). Eles argumentaram que as observações de White provavelmente não estavam relacionadas ao sexo dos animais. Isso foi apoiado pelo fato de que alguns dos espécimes da Formação Cutler tinham chevrons que apareceram pela primeira vez em sua quinta vértebra caudal. Embora fosse possível que o tamanho genital fosse variável entre os homens a ponto de impactar o esqueleto, a explicação mais provável era que as diferenças que White havia observado eram causadas por variação individual do esqueleto, divergência evolutiva ou algum outro fator não relacionado ao dimorfismo sexual. Da mesma forma, eles concordaram que as proporções do crânio apoiavam a proposta de Vaughn (1966) de que o dimorfismo estava presente nos fósseis de Seymouria , embora eles discordassem de como ele o relacionou ao sexo usando um fóssil que foi considerado "feminino" pelos critérios de White. A descoberta de seymouriamorphs larvais fossilizados mostrou que Seymouria provavelmente tinha um estágio larval aquático, desmentindo as hipóteses anteriores de que Seymouria botava ovos na terra.
Histologia e desenvolvimento
A evidência histológica de espécimes encontrados em Richards Spurs , Oklahoma , forneceu informações adicionais sobre a biologia de Seymouria . Um fêmur foi encontrado para ter uma estrutura interna caracterizada por uma matriz lamelar perfurada por numerosos canais plexiformes. As linhas de repouso de crescimento lento são indistintas e bem espaçadas, mas não há evidências de que o crescimento tenha cessado em qualquer momento durante o desenvolvimento ósseo. Como a maioria dos lissamfíbios , a cavidade medular é aberta e possui uma pequena quantidade de osso esponjoso. O desenvolvimento do osso esponjoso é ligeiramente superior ao de Acheloma (um anfíbio terrestre), mas é muito menos extenso do que os anfíbios aquáticos como Rhinesuchus e Trimerorhachis . As vértebras de Seymouria são mais robustas em forma em comparação com Discosauriscus , e têm uma pequena quantidade de cartilagem, apesar de uma grande quantidade de porosidade. Infere-se que a Seymouria sofreu metamorfose muito cedo na vida, provavelmente devido a estresses ambientais decorrentes da flutuação das estações chuvosa e seca.
Referências
links externos
- Uma fotografia do espécime "Tambach lovers", publicada pela conta de Mark MacDougall no Twitter
- Outra fotografia dos "amantes de Tambach", publicada pela "Geology Page"
- Uma fotografia do bloco Cutler Formation, publicada pela conta de Twitter de "mskvarla36"
- Documentário traduzido da DW sobre fósseis de Tambach, incluindo Seymouria
{kind=link}
{kind=link}
{kind=link}