
Carnotaurus -Carnotaurus
|
Carnotaurus |
|
|---|---|

|
|
| Molde esquelético montado no Museu Chlupáč em Praga | |
|
Classificação científica |
|
| Reino: | Animalia |
| Filo: | Chordata |
| Clade : | Dinosauria |
| Clade : | Saurischia |
| Clade : | Theropoda |
| Família: | † Abelisauridae |
| Clade : | † Furileusauria |
| Tribo: | † Carnotaurini |
| Gênero: |
† Carnotaurus Bonaparte , 1985 |
| Espécies: |
† C. sastrei
|
| Nome binomial | |
|
† Carnotaurus sastrei Bonaparte, 1985
|
|
Carnotaurus / ˌ k ɑr n oʊ t ɔr ə s / é um gênero de theropod dinossauro que viveu na América do Sul durante o Cretáceo período, provavelmente em algum momento entre 72 e 69,9 milhões de anos atrás. A única espécie é Carnotaurus sastrei . Conhecido por um único esqueleto bem preservado, é um dos terópodes mais conhecidos do hemisfério sul . O esqueleto, encontrado em 1984, foi descoberto na província de Chubut, na Argentina, a partir de rochas da Formação La Colonia . Carnotaurus é um membro derivado do Abelisauridae , um grupo de grandes terópodes que ocupou o grande nicho predatorial nas massas de terra do sul de Gondwana durante o final do Cretáceo . Dentro dos Abelisauridae, o gênero é frequentemente considerado um membro da Brachyrostra, um clado de formas de focinho curto restrito à América do Sul.
Carnotaurus era um predador bípede , de constituição leve , medindo 7,5 a 9 m (24,6 a 29,5 pés) de comprimento e pesando pelo menos 1,35 toneladas métricas (1,33 toneladas longas; 1,49 toneladas curtas). Como terópode, o carnotauro era altamente especializado e distinto. Tinha chifres grossos acima dos olhos, uma característica não vista em todos os outros dinossauros carnívoros , e um crânio muito profundo apoiado em um pescoço musculoso. O carnotauro foi caracterizado por pequenos membros anteriores vestigiais e membros posteriores longos e delgados. O esqueleto é preservado com impressões de pele extensas, mostrando um mosaico de pequenas escamas não sobrepostas com aproximadamente 5 mm de diâmetro. O mosaico foi interrompido por grandes saliências que revestiam as laterais do animal, e não há indícios de penas.
Os chifres característicos e o pescoço musculoso podem ter sido usados na luta contra co-específicos . De acordo com estudos separados, indivíduos rivais podem ter lutado uns contra os outros com golpes rápidos na cabeça, por empurrões lentos com a parte superior do crânio ou por bater uns nos outros de frente, usando seus chifres como amortecedores. Os hábitos alimentares do carnotauro permanecem obscuros: alguns estudos sugeriram que o animal era capaz de caçar presas muito grandes, como os saurópodes , enquanto outros estudos descobriram que ele se alimentava principalmente de animais relativamente pequenos. Sua cavidade cerebral sugere um olfato agudo, enquanto a audição e a visão eram menos desenvolvidas. O carnotauro provavelmente foi bem adaptado para correr e foi possivelmente um dos grandes terópodes mais rápidos.
Descoberta
O único esqueleto ( holótipo MACN-CH 894) foi descoberto em 1984 por uma expedição liderada pelo paleontólogo argentino José Bonaparte . Esta expedição também recuperou o peculiar saurópode espinhoso Amargasaurus . Foi a oitava expedição dentro do projeto "Vertebrados Terrestres Jurássicos e Cretáceos da América do Sul", que teve início em 1976 e foi patrocinado pela National Geographic Society . O esqueleto está bem preservado e articulado (ainda conectado), com apenas os dois terços posteriores da cauda, grande parte da perna e as patas traseiras sendo destruídas pelo intemperismo . O esqueleto pertencia a um indivíduo adulto, como indicam as suturas fundidas na caixa craniana . Foi encontrado deitado sobre o lado direito, mostrando uma pose típica de morte com o pescoço dobrado para trás sobre o tronco. Excepcionalmente, é preservado com impressões de pele extensas. Tendo em vista o significado dessas impressões, uma segunda expedição foi iniciada para reinvestigar o local da escavação original, levando à recuperação de várias manchas de pele adicionais. O crânio foi deformado durante a fossilização, com os ossos do focinho do lado esquerdo deslocados para frente em relação ao lado direito, os ossos nasais empurrados para cima e os pré - maxilares empurrados para trás nos ossos nasais . A deformação também exagerou a curvatura para cima da mandíbula superior. O focinho foi mais fortemente afetado pela deformação do que a parte posterior do crânio, possivelmente devido à maior rigidez desta. Na vista superior ou inferior, as mandíbulas superiores eram menos em forma de U do que as inferiores, resultando em uma aparente incompatibilidade. Este desencontro é o resultado da deformação atuando nas laterais, que afetou as mandíbulas superiores, mas não as inferiores, possivelmente devido à maior flexibilidade das articulações dentro destas.

O esqueleto foi coletado em uma fazenda chamada "Pocho Sastre" perto de Bajada Moreno, no departamento de Telsen, na província de Chubut , na Argentina. Por estar incrustado em uma grande concreção de hematita , um tipo de rocha muito dura, o preparo foi complicado e progrediu lentamente. Em 1985, Bonaparte publicou uma nota apresentando Carnotaurus sastrei como um novo gênero e espécie e descrevendo brevemente o crânio e a mandíbula. O nome genérico Carnotaurus é derivado do latim carno [carnis] ("carne") e taurus ("touro") e pode ser traduzido como "touro carnívoro", uma alusão aos chifres semelhantes aos de touro do animal. O nome específico sastrei homenageia Angel Sastre, o proprietário da fazenda onde o esqueleto foi encontrado. Uma descrição abrangente de todo o esqueleto ocorreu em 1990. Depois do Abelisaurus , o Carnotaurus foi o segundo membro da família Abelisauridae a ser descoberto. Durante anos, foi de longe o membro mais bem compreendido de sua família e também o terópode mais bem compreendido do hemisfério sul . Não foi até o século 21 que abelisaurídeos bem preservados semelhantes foram descritos, incluindo Aucasaurus , Majungasaurus e Skorpiovenator , permitindo aos cientistas reavaliar certos aspectos da anatomia do Carnotaurus . O esqueleto do holótipo está exposto no Museu Argentino de Ciências Naturais , Bernardino Rivadavia ; réplicas podem ser vistas neste e em outros museus ao redor do mundo. Os escultores Stephen e Sylvia Czerkas fabricaram uma escultura em tamanho real de Carnotaurus que estava anteriormente em exibição no Museu de História Natural do Condado de Los Angeles . Esta escultura, encomendada pelo museu em meados da década de 1980, é provavelmente a primeira restauração de vida de um terópode mostrando uma pele precisa.
Descrição

O carnotauro era um predador grande, mas de constituição leve. O único indivíduo conhecido tinha cerca de 7,5-9 m (24,6-29,5 pés) de comprimento, tornando o Carnotaurus um dos maiores abelisaurídeos. Ekrixinatosaurus e possivelmente Abelisaurus , que são altamente incompletos, podem ter sido semelhantes ou maiores em tamanho. Um estudo de 2016 descobriu que apenas o Pycnonemosaurus , a 8,9 m (29,2 pés), era mais longo do que o Carnotaurus ; foi estimado em 7,8 m (25,6 pés). Sua massa é estimada em 1.350 kg (1,33 toneladas longas; 1,49 toneladas curtas), 1.500 kg (1,5 toneladas longas; 1,7 toneladas curtas), 2.000 kg (2,0 toneladas longas; 2,2 toneladas curtas), 2.100 kg (2,1 toneladas longas; 2,3 toneladas curtas) e 1.306-1.743 kg (1.285-1.715 toneladas longas; 1.440-1.921 toneladas curtas) em estudos separados que usaram diferentes métodos de estimativa. O carnotauro era um terópode altamente especializado , visto especialmente nas características do crânio , das vértebras e dos membros anteriores. A pelve e os membros posteriores, por outro lado, permaneceram relativamente conservadores, assemelhando-se aos do Ceratossauro mais basal . Tanto a pélvis quanto os membros posteriores eram longos e delgados. O fêmur esquerdo (osso da coxa) do indivíduo mede 103 cm de comprimento, mas apresenta diâmetro médio de apenas 11 cm.
Crânio

O crânio, medindo 59,6 cm (23,5 polegadas) de comprimento, era proporcionalmente mais curto e mais profundo do que em qualquer outro grande dinossauro carnívoro. O focinho era moderadamente largo, não tão estreito como visto em terópodes mais basais como o ceratossauro , e as mandíbulas eram curvadas para cima. Um par de chifres proeminentes se projetava obliquamente acima dos olhos. Esses chifres, formados pelos ossos frontais , eram grossos e em forma de cone, internamente sólidos, um tanto achatados verticalmente na seção transversal e mediam 15 cm (5,9 pol.) De comprimento. Bonaparte, em 1990, sugeriu que esses chifres provavelmente teriam formado os núcleos ósseos de bainhas queratinosas muito mais longas. Mauricio Cerroni e colegas, em 2020, concordaram que os chifres suportavam bainhas queratinosas, mas argumentaram que essas bainhas não seriam muito mais longas que os núcleos ósseos.
Como em outros dinossauros, o crânio foi perfurado por seis aberturas de crânio principais em cada lado. A mais frontal dessas aberturas, a naris externa (narina óssea), era sub-retangular e direcionada para os lados e para a frente, mas não era inclinada em vista lateral como em alguns outros ceratossauros como o Ceratossauro . Essa abertura era formada apenas pelo nasal e pré-maxilar, enquanto em alguns ceratossauros relacionados a maxila também contribuía para essa abertura. Entre a narina óssea e a órbita (abertura do olho) ficava a fenestra antorbital . No Carnotaurus , essa abertura era mais alta do que longa, enquanto era mais longa do que alta em formas relacionadas, como Skorpiovenator e Majungasaurus . A janela antorbital era delimitada por uma depressão maior, a fossa antorbital , que era formada por partes rebaixadas da maxila na frente e as lacrimais atrás. Como em todos os abelissaurídeos, essa depressão era pequena no carnotauro . O canto frontal inferior da fossa antorbital continha uma abertura menor, a fenestra promaxilar , que conduzia a uma cavidade cheia de ar dentro da maxila. O olho estava situado na parte superior da órbita em forma de buraco de fechadura. Esta parte superior era proporcionalmente pequena e subcircular, e separada da parte inferior da órbita pelo osso pós -orbital que se projetava para frente . Estava ligeiramente girado para a frente, provavelmente permitindo algum grau de visão binocular . A forma semelhante a um buraco de fechadura da órbita foi possivelmente relacionada ao acentuado encurtamento do crânio e também é encontrada em abelisaurídeos de focinho curto. Como em todos os abelissaurídeos, o osso frontal (no teto do crânio, entre os olhos) foi excluído da órbita. Atrás da órbita havia duas aberturas, a janela infratemporal na lateral e a janela supratemporal no topo do crânio. A fenestra infratemporal era alta, curta e em formato de rim, enquanto a fenestra supratemporal era curta e quadrada. Outra abertura, a fenestra mandibular , estava localizada na mandíbula - no Carnotaurus , essa abertura era comparativamente grande.

Em cada lado das maxilas superiores havia quatro pré-maxilar e doze maxilares dentes, enquanto que as maxilas inferiores foram equipados com quinze dentário dentes de cada lado. Os dentes foram descritos como longos e delgados, ao contrário dos dentes muito curtos vistos em outros abelisaurídeos. No entanto, Cerroni e colegas, em sua descrição do crânio em 2020, afirmaram que todos os dentes erupcionados foram gravemente danificados durante a escavação e foram posteriormente reconstruídos com gesso (Bonaparte, em 1990, observou apenas que alguns dentes da mandíbula inferior haviam sido fragmentados). Informações confiáveis sobre a forma dos dentes são, portanto, limitadas aos dentes substitutos e às raízes dos dentes que ainda estão contidos na mandíbula e podem ser estudadas por meio de imagens de TC. Os dentes substitutos tinham coroas baixas e achatadas , espaçados próximos e inclinados para frente em aproximadamente 45 °. Em sua descrição de 1990, Bonaparte observou que a mandíbula inferior era rasa e fracamente construída, com o dentário (o osso anterior da mandíbula) conectado aos ossos posteriores da mandíbula por apenas dois pontos de contato; isso contrasta com o crânio de aparência robusta. Em vez disso, Cerroni e seus colegas encontraram conexões múltiplas, mas frouxas, entre o dentário e os ossos posteriores da mandíbula. Essa articulação, portanto, era muito flexível, mas não necessariamente fraca. A margem inferior do dentário era convexa, ao passo que era reta no Majungassauro .

A mandíbula inferior foi encontrada com ossos hióide ossificados , na posição em que estariam se o animal estivesse vivo. Esses ossos delgados, que sustentam a musculatura da língua e vários outros músculos, raramente são encontrados nos dinossauros porque geralmente são cartilaginosos e não estão conectados a outros ossos e, portanto, se perdem facilmente. No Carnotaurus , três ossos hióide são preservados: um par de ceratobranquiais curvos em forma de bastonete que se articulam com um único elemento trapezoidal, o basi-hial. Carnotaurus é o único terópode não aviário conhecido do qual um basihyal é conhecido. A parte posterior do crânio tinha câmaras bem desenvolvidas e cheias de ar ao redor da caixa craniana, como em outros abelisaurídeos. Dois sistemas separados de câmaras estavam presentes, o sistema paratimpânico, que estava conectado à cavidade do ouvido médio , bem como as câmaras resultantes do crescimento dos sacos aéreos do pescoço.
Várias autapomorfias (características distintivas) podem ser encontradas no crânio, incluindo o par de chifres e o crânio muito curto e profundo. A maxila tinha escavações acima da fenestra promaxilar, que teria sido escavada pelo seio aéreo antorbital (passagens de ar no focinho). O ducto nasolacrimal, que transporta o fluido ocular, sai na superfície medial (interna) do lacrimal por um canal de função incerta. Outras autapomorfias propostas incluem uma escavação profunda e longa preenchida com ar no quadrato e uma depressão alongada no pterigóide do palato .
Vértebras

A coluna vertebral consistia em dez cervicais (pescoço), doze dorsais , seis sacrais fundidas e um número desconhecido de vértebras caudais (cauda). O pescoço era quase reto, em vez de ter a curva em S vista em outros terópodes, e também anormalmente largo, especialmente em direção à sua base. O topo da coluna vertebral do pescoço apresentava uma fileira dupla de processos ósseos aumentados e direcionados para cima, chamados epipófises , criando uma depressão lisa na parte superior das vértebras do pescoço. Esses processos eram os pontos mais altos da espinha, elevando-se acima dos processos espinhosos incomumente baixos . As epipófises provavelmente forneciam áreas de inserção para uma musculatura do pescoço marcadamente forte. Uma fileira dupla semelhante também estava presente na cauda, formada por costelas caudais altamente modificadas , em vista frontal projetando-se para cima em forma de V, seus lados internos criando uma superfície superior lisa e plana das vértebras da cauda dianteira. A extremidade de cada costela caudal era fornecida com uma expansão em forma de gancho projetada para a frente que se conectava à costela caudal da vértebra anterior.
Membros anteriores


Os membros anteriores eram proporcionalmente mais curtos do que em qualquer outro grande dinossauro carnívoro, incluindo o tiranossauro . O antebraço tinha apenas um quarto do tamanho do braço. Não havia carpalia na mão, de modo que os metacarpos se articulavam diretamente com o antebraço. A mão apresentava quatro dígitos básicos, embora aparentemente apenas os dois do meio terminassem em ossos dos dedos, enquanto o quarto consistia em um único metacarpo semelhante a uma tala que pode ter representado um "esporão" externo. Os próprios dedos estavam fundidos e imóveis, e podem não ter garras. Carnotaurus diferia de todos os outros abelisaurídeos por ter membros anteriores proporcionalmente mais curtos e mais robustos e por ter o quarto metacarpo semelhante a uma tala como o osso mais longo da mão. Um estudo de 2009 sugere que os braços eram vestigiais em abelisaurídeos, porque as fibras nervosas responsáveis pela transmissão do estímulo foram reduzidas a uma extensão vista nos emas e kiwis de hoje , que também têm membros anteriores vestigiais.
Pele

Carnotaurus foi o primeiro dinossauro terópode descoberto com um número significativo de impressões de pele fóssil . Essas impressões, encontradas abaixo do lado direito do esqueleto, vêm de diferentes partes do corpo, incluindo a mandíbula, a parte frontal do pescoço, a cintura escapular e a caixa torácica . A maior mancha de pele corresponde à parte anterior da cauda. Originalmente, o lado direito do crânio também era coberto por grandes manchas de pele - isso não foi reconhecido quando o crânio foi preparado e essas manchas foram destruídas acidentalmente. No entanto, a textura da superfície de vários ossos do crânio permite inferências sobre sua provável cobertura. Uma superfície montanhosa com sulcos, fossas e pequenas aberturas é encontrada nas laterais e na frente do focinho e indica uma cobertura escamosa, possivelmente com escamas planas como nos crocodilianos de hoje. O topo do focinho foi esculpido com numerosos pequenos orifícios e pontas - esta textura pode provavelmente ser correlacionada com uma almofada cornificada (cobertura córnea). Essa almofada também ocorreu no Majungasaurus, mas estava ausente no Abelisaurus e Rugops . Provavelmente, uma fileira de grandes escamas circundava o olho, conforme indicado por uma superfície ondulada com sulcos longitudinais nos ossos lacrimal e pós-orbital.
A pele foi construída a partir de um mosaico de escalas poligonais não sobrepostas medindo aproximadamente 5–12 mm (0,20–0,47 pol.) De diâmetro. Este mosaico foi dividido por ranhuras finas e paralelas. A escala era semelhante em diferentes partes do corpo, com exceção da cabeça, que aparentemente apresentava um padrão diferente e irregular de escamas. Não há evidência de penas. Estruturas maiores em forma de colisão foram distribuídas nas laterais do pescoço, costas e cauda em fileiras irregulares. Essas saliências tinham de 4 a 5 cm (1,6 a 2,0 pol.) De diâmetro e até 5 cm (2,0 pol.) De altura e geralmente exibiam uma crista baixa na linha média. Eles foram colocados 8 a 10 cm (3,1 a 3,9 pol.) Separados um do outro e tornaram-se maiores em direção ao topo do animal. As saliências provavelmente representam escamas - agrupamentos de escamas condensadas - semelhantes às vistas no babado macio ao longo da linha média do corpo em dinossauros hadrossaurídeos ("bico de pato"). Essas estruturas não continham osso. Stephen Czerkas (1997) sugeriu que essas estruturas podem ter protegido os lados do animal enquanto lutavam contra membros da mesma espécie ( co-específicos ) e outros terópodes, argumentando que estruturas semelhantes podem ser encontradas no pescoço da iguana moderna, onde fornecem proteção limitada em combate .
Estudos mais recentes da pele do Carnotaurus publicados em 2021 sugerem que as representações anteriores das escamas no corpo são imprecisas e as escalas de características maiores foram distribuídas aleatoriamente ao longo do corpo, não distribuídas em linhas discretas como em representações artísticas e ilustrações mais antigas. Também não há nenhum sinal de variação progressiva de tamanho nas escalas de recursos ao longo de diferentes áreas ao longo do corpo. As escalas basais do Carnotaurus eram, em comparação, altamente variáveis, variando em tamanho de pequeno e alongado a grande e poligonal, e de circular a lenticular nas regiões torácica, escapular e cauda, respectivamente. Essa diferenciação de escala pode estar relacionada à regulação do calor corporal e à liberação do excesso de calor por meio da termorregulação devido ao seu grande tamanho corporal e estilo de vida ativo.
Classificação


Carnotaurus é um dos gêneros mais conhecidos de Abelisauridae , uma família de grandes terópodes restrita ao antigo supercontinente sul Gondwana . Os abelisaurídeos foram os predadores dominantes no Cretáceo Superior de Gondwana, substituindo os carcarodontossaurídeos e ocupando o nicho ecológico preenchido pelos tiranossaurídeos nos continentes do norte. Vários traços notáveis que evoluíram dentro desta família, incluindo encurtamento do crânio e braços, bem como peculiaridades nas vértebras cervicais e caudais, eram mais pronunciados no Carnotaurus do que em qualquer outro abelisaurídeo.
Embora as relações dentro dos Abelisauridae sejam debatidas, o Carnotaurus é consistentemente mostrado como um dos membros mais derivados da família por análises cladísticas. Seu parente mais próximo pode ter sido Aucasaurus ou Majungasaurus . Em contraste, uma revisão de 2008 sugeriu que o Carnotaurus não estava intimamente relacionado a nenhum dos gêneros e, em vez disso, propôs Ilokelesia como seu táxon irmão . Juan Canale e colegas, em 2009, erigiram o novo clado Brachyrostra para incluir Carnotaurus, mas não Majungasaurus ; essa classificação foi seguida por uma série de estudos desde então.
Carnotaurus é homônimo para dois subgrupos de Abelisauridae: o Carnotaurinae e o Carnotaurini . Os paleontólogos não aceitam universalmente esses grupos. O Carnotaurinae foi definido para incluir todos os abelisaurídeos derivados com a exclusão do Abelisaurus , que é considerado um membro basal na maioria dos estudos. No entanto, uma revisão de 2008 sugeriu que o Abelisaurus era um abelisaurídeo derivado. Carnotaurini foi proposto para nomear o clado formado por Carnotaurus e Aucasaurus ; somente os paleontólogos que consideram o Aucasaurus como o parente mais próximo do Carnotaurus usam esse grupo.
Abaixo está um cladograma publicado por Canale e colegas em 2009.
| Carnotaurinae |
|
||||||||||||||||||||||||||||||
Paleobiologia
Função dos chifres

Carnotaurus é o único animal carnívoro bípede conhecido com um par de chifres no osso frontal. O uso desses chifres não está totalmente claro. Várias interpretações giraram em torno do uso na luta contra membros da mesma espécie ou no abate de presas, embora também seja possível um uso em exibição para cortejo ou reconhecimento de membros da mesma espécie.
Greg Paul (1988) propôs que os chifres eram armas de cabeçada e que a pequena orbita teria minimizado a possibilidade de ferir os olhos durante a luta. Gerardo Mazzetta e colegas (1998) sugeriram que o Carnotaurus usava seus chifres de forma semelhante aos carneiros. Eles calcularam que a musculatura do pescoço era forte o suficiente para absorver a força de dois indivíduos colidindo com suas cabeças frontalmente a uma velocidade de 5,7 m / s cada. Fernando Novas (2009) interpretou várias características do esqueleto como adaptações para desferir golpes com a cabeça. Ele sugeriu que o encurtamento do crânio poderia ter tornado os movimentos da cabeça mais rápidos, reduzindo o momento de inércia , enquanto o pescoço musculoso teria permitido fortes golpes de cabeça. Ele também notou uma maior rigidez e força da coluna vertebral que pode ter evoluído para suportar choques conduzidos pela cabeça e pescoço.
Outros estudos sugerem que o carnotauro rival não dava golpes rápidos na cabeça, mas empurrava lentamente um contra o outro com a parte superior do crânio. Mazzetta e colegas, em 2009, argumentaram que os chifres podem ter sido um dispositivo para a distribuição de forças de compressão sem danos ao cérebro. Isso é sustentado pelos lados superiores achatados dos chifres, pelos ossos fortemente fundidos do topo do crânio e pela incapacidade do crânio de sobreviver a golpes rápidos na cabeça. Rafael Delcourt, em 2018, sugeriu que os chifres poderiam ter sido usados tanto em cabeçadas e empurrões lentos, como na iguana marinha moderna , quanto em golpes no pescoço e flancos do oponente, como na girafa moderna . A última possibilidade havia sido proposta anteriormente para o Majungasaurus relacionado em um documento de conferência de 2011.
Gerardo Mazzetta e colegas (1998) propõem que os chifres também podem ter sido usados para ferir ou matar pequenas presas. Embora os núcleos dos chifres sejam rombos, eles podem ter tido uma forma semelhante aos chifres bovinos modernos se houvesse uma cobertura de queratina . No entanto, este seria o único exemplo relatado de chifres sendo usados como armas de caça em animais.
Função mandibular e dieta

As análises da estrutura da mandíbula do Carnotaurus por Mazzetta e colegas, em 1998, 2004 e 2009, sugerem que o animal era capaz de mordidas rápidas, mas não fortes. Mordidas rápidas são mais importantes do que mordidas fortes ao capturar pequenas presas , como mostrado por estudos de crocodilos modernos . Esses pesquisadores também notaram um alto grau de flexibilidade ( cinesia ) dentro do crânio e especialmente na mandíbula, um tanto semelhante às cobras modernas . A elasticidade da mandíbula teria permitido que o carnotauro engolisse pequenas presas inteiras. Além disso, a parte frontal da mandíbula inferior era articulada e, portanto, capaz de se mover para cima e para baixo. Quando pressionados para baixo, os dentes teriam se projetado para a frente, permitindo que o carnotauro espetasse pequenas presas; quando os dentes estavam curvados para cima, os dentes agora projetados para trás teriam impedido a presa capturada de escapar. Mazzetta e seus colegas também descobriram que o crânio foi capaz de resistir às forças que aparecem ao puxar grandes presas. O carnotauro pode, portanto, ter se alimentado principalmente de presas relativamente pequenas, mas também foi capaz de caçar grandes dinossauros. Em 2009, Mazzetta e colegas estimaram uma força de mordida de cerca de 3.341 newtons.
Essa interpretação foi questionada por François Therrien e colegas (2005), que descobriram que a força de mordida do Carnotaurus era o dobro do jacaré americano , que pode ter a mordida mais forte de qualquer tetrápode vivo . Esses pesquisadores também notaram analogias com os dragões de Komodo modernos : a resistência à flexão da mandíbula inferior diminui em direção à ponta linearmente, indicando que as mandíbulas não eram adequadas para captura de presas pequenas com alta precisão, mas para fazer cortes para enfraquecer presas grandes. Como consequência, de acordo com este estudo, o Carnotaurus deve ter predado principalmente animais de grande porte, possivelmente por emboscada. Cerroni e colegas, em 2020, argumentaram que a flexibilidade estava restrita à mandíbula, enquanto o teto do crânio engrossado e a ossificação de várias articulações cranianas sugerem que o crânio não tinha ou tinha apenas pouca cinesia.
Robert Bakker (1998) descobriu que o Carnotaurus se alimentava principalmente de presas muito grandes, especialmente saurópodes . Como ele observou, várias adaptações do crânio - o focinho curto, os dentes relativamente pequenos e a parte posterior forte do crânio ( occipital ) - evoluíram independentemente no Allosaurus . Essas características sugerem que a mandíbula superior era usada como uma clava serrilhada para infligir feridas; grandes saurópodes teriam sido enfraquecidos por ataques repetidos.
Locomoção


Mazzetta e colegas (1998, 1999) presumiram que o Carnotaurus era um corredor veloz, argumentando que o osso da coxa foi adaptado para resistir a momentos de alta flexão durante a corrida. A capacidade da perna de um animal de resistir a essas forças limita sua velocidade máxima. As adaptações de corrida do carnotauro teriam sido melhores do que as de um humano, embora não tão boas quanto as de um avestruz . Os cientistas calculam que o Carnotaurus teve uma velocidade máxima de até 48-56 km (30-35 mi) por hora.
Nos dinossauros, o músculo locomotor mais importante estava localizado na cauda. Esse músculo, chamado caudofemoral , se liga ao quarto trocânter , uma crista proeminente no osso da coxa, e puxa o osso da coxa para trás quando contraído. Scott People e Phil Currie (2011) observam que nas vértebras da cauda do Carnotaurus , as costelas caudais não se projetavam horizontalmente ("em forma de T"), mas eram anguladas contra o eixo vertical das vértebras, formando um "V". Isso teria fornecido espaço adicional para um músculo caudofemoral maior do que em qualquer outro terópode - a massa muscular foi calculada em 111 a 137 quilogramas (245 a 302 lb) por perna. Portanto, o Carnotaurus poderia ter sido um dos grandes terópodes mais rápidos. Enquanto o músculo caudofemoral estava aumentado, os músculos epaxiais situados acima das costelas caudais seriam proporcionalmente menores. Esses músculos, chamados de músculo longissimus e espinhal , eram responsáveis pelo movimento e estabilidade da cauda. Para manter a estabilidade da cauda apesar da redução desses músculos, as costelas caudais suportam processos de projeção para a frente, interligando as vértebras entre si e com a pelve, enrijecendo a cauda. Como consequência, a capacidade de fazer curvas fechadas teria diminuído, porque o quadril e a cauda precisavam ser virados simultaneamente, ao contrário de outros terópodes.
Cérebro e sentidos
Cerroni e Paulina-Carabajal, em 2019, usaram uma tomografia computadorizada para estudar a cavidade endocraniana que continha o cérebro. O volume da cavidade endocraniana era de 168,8 cm³, embora o cérebro só tivesse preenchido uma fração desse espaço. Os autores usaram duas estimativas de tamanho do cérebro diferentes, assumindo um tamanho do cérebro de 50% e 37% da cavidade endocraniana, respectivamente. Isso resulta em um quociente de encefalização réptil (uma medida de inteligência) maior do que o do Majungasaurus aparentado, mas menor do que nos tiranossaurídeos . A glândula pineal , que produz hormônios , pode ter sido menor do que em outros abelisaurídeos, conforme indicado por uma expansão dural baixa - um espaço no topo do prosencéfalo no qual a glândula pineal está localizada.
Os bulbos olfativos , que albergavam o sentido do olfato, eram grandes, enquanto os lobos ópticos , responsáveis pela visão, eram relativamente pequenos. Isso indica que o sentido do olfato pode ter sido melhor desenvolvido do que o sentido da visão, enquanto o oposto é o caso nos pássaros modernos. A extremidade frontal dos tratos olfativos e bulbos eram curvados para baixo, uma característica compartilhada apenas pelo Indosaurus ; em outros abelisaurídeos, essas estruturas eram orientadas horizontalmente. Conforme a hipótese de Cerroni e Paulina-Carabajal, essa curvatura para baixo, juntamente com o grande tamanho dos bulbos, pode indicar que o Carnotaurus confiava mais no olfato do que outros abelisaurídeos. O flóculo , um lobo cerebral que se acredita estar relacionado à estabilização do olhar (coordenação entre os olhos e o corpo), era grande no carnotauro e em outros abelisaurídeos sul-americanos. Isso pode indicar que essas formas freqüentemente usam movimentos rápidos da cabeça e do corpo. A audição pode ter sido pouco desenvolvida em Carnotaurus e outros abelisaurídeos, como indicado pela lagena curta do ouvido interno . A faixa de audição foi estimada em menos de 3 kHz.
Idade e paleoambiente

Originalmente, as rochas nas quais o Carnotaurus foi encontrado foram atribuídas à parte superior da Formação Gorro Frigio , que foi considerada ter aproximadamente 100 milhões de anos ( estágio Albiano ou Cenomaniano ). Mais tarde, eles foram percebidos como pertencentes à formação La Colonia , muito mais jovem , datando dos estágios Campaniano e Maastrichtiano (83,6 a 66 milhões de anos atrás). Novas, em um livro de 2009, deu um intervalo de tempo mais estreito de 72 a 69,9 milhões de anos atrás ( estágio Maastrichtiano inferior ). Carnotaurus, portanto, foi o último abelisaurídeo sul-americano conhecido. No final do Cretáceo, a América do Sul já estava isolada da África e da América do Norte.
A Formação La Colonia está exposta na encosta sul do Maciço Patagônico Norte . A maioria dos fósseis de vertebrados, incluindo o Carnotaurus , vem da seção intermediária da formação (chamada de associação de fácies intermediária ). Esta parte provavelmente representa os depósitos de um ambiente de estuários , planícies de marés ou planícies costeiras . O clima teria sido sazonal com períodos secos e úmidos. Os vertebrados mais comuns coletados incluem peixes pulmonados ceratodontídeos , tartarugas, crocodilos, plesiossauros , dinossauros, lagartos, cobras e mamíferos. Algumas das cobras encontradas pertencem às famílias Boidae e Madtsoidae, como Alamitophis argentinus . As tartarugas são representadas por pelo menos cinco táxons , quatro de Chelidae ( Pleurodira ) e um de Meiolaniidae ( Cryptodira ). Entre os répteis marinhos está o plesiossauro Sulcusuchus erraini da família Polycotylidae . Os mamíferos são representados por Reigitherium bunodontum , que foi considerado o primeiro registro de um docodonte sul-americano , e Argentodites coloniensis , possivelmente de Multituberculata . Em 2011, foi anunciada a descoberta de um novo pássaro enantiornithine da Formação La Colonia.
Veja também
Notas
Referências
links externos
- A mordida do Carnotaurus na Universidad Nacional de Mar del Plata. (em espanhol)
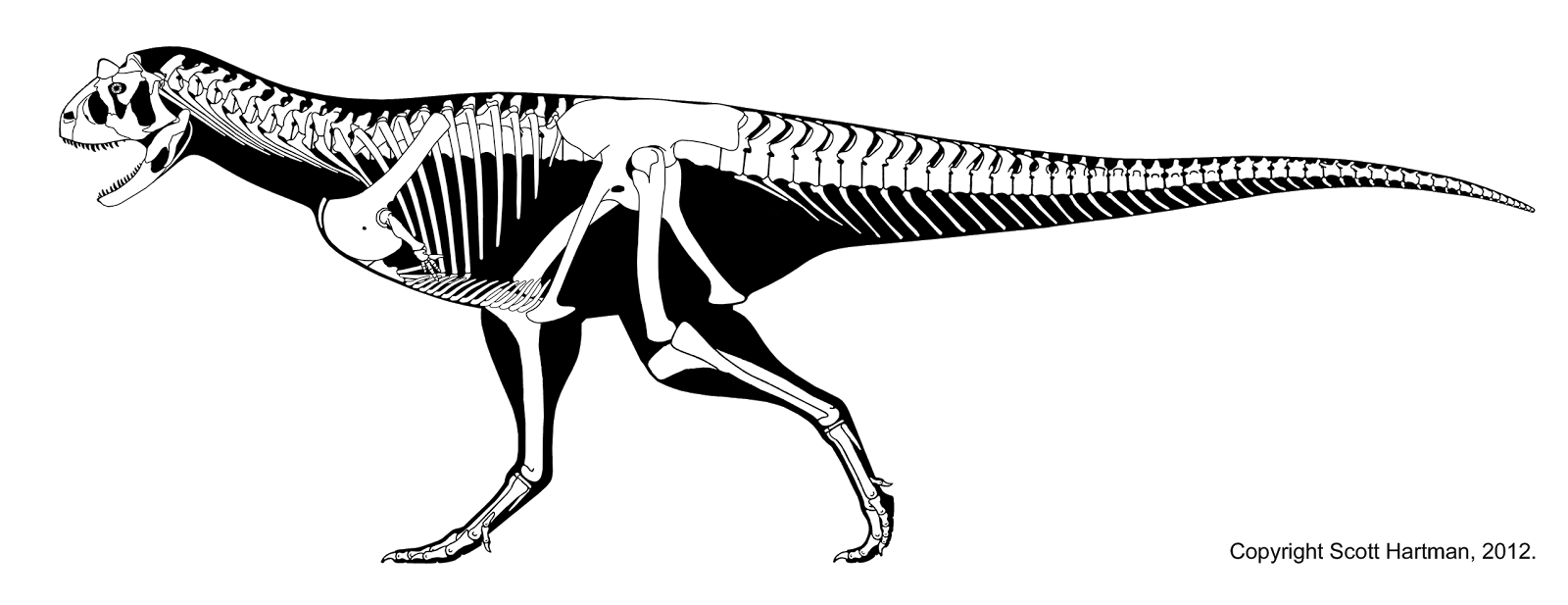
- Reconstrução do esqueleto por Scott Hartman
{kind=link}